 ORIGINAL RESEARCH
ORIGINAL RESEARCH
Effect of dietary chromium tripicolinate on productivity of sows under commercial conditions
Chad D. Hagen, PhD; Merlin D. Lindemann, PhD; Ken W. Purser, MS, PhD
Hagen CD, Lindemann MD, Purser KW. Effect of dietary chromium tripicolinate on productivity of sows under commercial conditions. Swine Health Prod. 2000;8(2):59-63. This article is also available in PDF format.
CDH: Iowa Select Farms; Reprint requests to MDL: University of Kentucky; 611 W.P. Garrigus Building, Lexington, Kentucky 40546-0215; email: mdlind1@pop.uky.edu; KWP: Prince Agri Products, Inc.
Summary
Objective: To further examine and clarify the effect of supplemental chromium (Cr) from chromium tripicolinate on productivity of sows (especially litter size) under commercial conditions.
Methods: Based on reproductive performance during a 3-month "pretest" period, 12 4000-sow units were assigned to one of two dietary treatments: either to a "+Cr" group that received 200 ppb supplemental Cr from chromium tripicolinate, or to a "Control" group that received a standard commercial ration without supplemental Cr. After a 6-month "loading" period, reproductive performance was evaluated over a 12-month "test" period.
Results: Dietary supplementation of 200 ppb Cr from chromium tripicolinate had a positive impact on sows bred by 7 days postweaning, number of pigs born alive and weaned, and sow mortality.
Implications: The use of a biologically available form of Cr can positively affect total sow productivity and allow greater animal output with no additional fixed capital inputs. This has the potential to improve total enterprise profitability.
 Keywords: chromium,
sows, litter size, sow productivity
Keywords: chromium,
sows, litter size, sow productivity
Received: February 26, 1999
Accepted: January 11, 2000
Sow productivity is a key factor in modern pork production. Its direct effect on animal numbers and ability to meet production volume objectives, combined with its indirect effect on cost of production through overhead allocation, make it a measure closely scrutinized at all levels of management. The two principle components of sow productivity are litter size and farrowing rate.
Recent research using supplemental chromium tripicolinate (CrP) has demonstrated substantial improvements in sow productivity. Studies1,2 have shown that CrP is associated with increased litter size. Australian research3,4 in a series of four experiments with > 4000 sows, demonstrated improved farrowing rates and more sows bred within 7 days of weaning in sows fed supplemental CrP. Based on these data, Campbell and Purser4 concluded that the use of CrP in breeding herd diets would improve sow productivity by 0.4-1.0 pigs per sow per year.
The objective of the present study was to further examine and clarify the effect of supplemental CrP on productivity of sows in commercial conditions, particularly on litter size.
Materials and methods
Facilities and management
Twelve 4000-sow units under the same ownership and located in the same state were selected for the study. All units were under the same general management and were served by the same feed mill. Breeding stock, facility design, equipment, and animal management practices were uniform across all units. Any changes that occurred during this study with respect to any of these factors were kept uniform across all experimental units.
During gestation, sows were housed in individual crates. On approximately day 112 of gestation, sows were moved to farrowing rooms and housed in farrowing crates. Pigs were weaned at 16-17 days of age and sows were bred on the first heat after weaning. At the beginning of the trial, the first service was a natural service with second and third services by artificial insemination (AI) in keeping with standard site practice. A transition to 100% AI was made during the course of the trial at a similar time at all sow units. Sows were predominantly PIC Camborough 15 at the start of the study with a transition to PIC Camborough 22 by conclusion of the study. Replacement gilts were moved to the gestation building in the sow units at about 68 kg (150 lb) bodyweight (BW) and were bred by standard practices at about 127 kg (280 lb).
Experimental design
For allotment to one of two dietary treatment groups in September 1996, the 12 4000-sow units were ranked from highest to lowest based on pigs weaned per mated sow per year (PWMSY) as recorded on the PigCHAMP(R) database during a 3-month "pretest" period. Beginning with the highest-ranking pair, one unit was then randomly allotted within that pair to one of the two dietary treatments, either:
- a "+Cr" treatment, in which supplemental CrP was added to provide 200 ppb Cr to the breeding, gestation, and lactation diets. CrP-supplemented diets were obtained by replacing 0.05% ground limestone in the control diets with an equal amount of CHROMAX(TM) brand CrP premix (0.04% Cr, Prince Agri Products, Inc.; Quincy, Illinois); or
- a "Control" treatment, which received no supplemental CrP.
Experimental diets were corn-soybean meal based with added wheat middlings in gestation and added fat in lactation. Actual ingredient use varied with the economics of the ingredient market. Lysine concentrations were 0.55% for gestation, 0.95% for lactation, and 0.85% for breeding. In a procedure similar to that previously described for wheat middlings and fat, the use of lysine HCl varied depending on economics. In no case did the concentration of lysine HCl exceed 0.10%. Chlortetracycline was added at 220 ppm (200 g per ton) to the breeding and lactation diets. The lactation diet was fed in the farrowing room and the breeding diet was fed after weaning through the end of the third service and to nongestating gilts. The gestation diet was fed at all other times.
After allotment, there was a 1-month "startup" period to allow each unit to begin use of the appropriate experimental diets. The start-up period was followed by a 6-month "loading" period to allow sows and replacement gilts adequate time to build body chromium stores and to assure that reproductive data obtained was only for sows that had received the dietary treatment for the entire gestational period. Finally, sow performance was evaluated over the 12-month "test" period by evaluating the PigCHAMP(R) records of the twelve sow units.
Statistical analysis
Analysis of covariance5 was performed using the GLM procedure of SAS (SAS; Cary, North Carolina) with the pre-assignment PWMSY as the covariate in the analysis. The continuous function variable of PWMSY ranged from 19.5-24.9. Values for each response parameter tested were those that PigCHAMP(R) calculated for the 1-year test period for each of the 12 units. These values were further divided for each unit into four parity groupings (parities one, two, three, and greater than three) to assess the potential for a unique response at different parities. Because there were no dietary treatmentxparity interactions, the final model included only dietary treatment as the single factor along with the covariate.
Problems with porcine reproductive and respiratory syndrome virus (PRRSV) were experienced during the "test" period in the sow units used in this study. We conducted an ANOVA on abortion data to evaluate the possibility that PRRSV may have contributed to a bias in the results. Although the number of abortions did not differ statistically (P >.20) between treatments, we conducted a test for potential outliers.6,7 The statistical examination of the data to assess the impact of the PRRSV outbreak on abortions by unit by month revealed some months in sow units on each treatment that had higher abortions than normal. The data set was temporarily adjusted to account for abortion occurrences in these months and the analyses for farrowing rate conducted again using analysis of covariance.
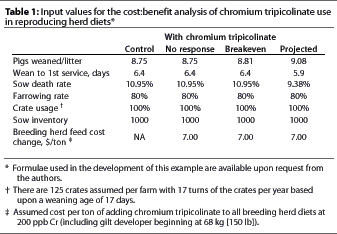
Cost:benefit evaluation
To put the observed biological results into an economic perspective, several cost:benefit scenarios were modeled using the total farm economic model developed for a Lotus spreadsheet (Compass Record Keeping System 1983, Hog Cashflow; Ralston Purina, Inc.; St. Louis, Missouri) (Table 1). To project potential income over feed cost using these cost and price assumptions, three different scenarios beyond the base "Control" scenario were calculated:
{kind=link}
- a "No response" scenario modelling the effect of adding supplemental CrP to all breeding herd diets in a hypothetical 1000-sow herd with no biological response;
- a "Breakeven" scenario in which litter size was adjusted to the point closest to breakeven ($0 profit) with supplemental CrP; and
- a "Projected" scenario modelling improvements in performance similar to those we observed in the present study in sows in the +Cr group.
Common assumptions used in this evaluation were of a 1000-sow farm with 125 farrowing crates that was weaning pigs at 17 days of age. Market hogs were sold at 118 kg (260 lb). Costs used for corn and 44% protein soybean meal and prices received for market hogs and cull sows were the 10-year historical averages.8 Nonfeed costs were set at zero based upon the assumption that they would not be affected by the changes observed in these data. Common industry values were used for other input parameters (Table 1) and were kept constant.
Results
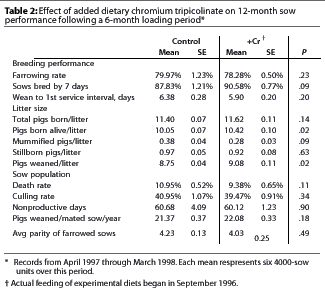
The allotment procedure resulted in an equalization of pigs born alive per litter (the primary response of interest) during the pretest period (10.37 for Controls versus 10.37 for the +Cr group). Additionally, the evaluation of other reproductive responses demonstrated that there were no measured differences between treatments (P >.10) during the pretest or the loading period.
Sows in the +Cr group had a numerically (although nonsignificant, P =.23) lower farrowing rate than Control sows (Table 2).
{kind=link}
After adjusting for the PRRSV-caused abortions, farrowing rate was virtually identical in the two treatment groups (80.00% versus 79.89%). Additionally, the adjustment did not alter the difference or statistical significance of any other response parameters, such as litter size, during the test period. Therefore, the nonadjusted values are shown in Table 2.
The use of supplemental CrP did improve aspects of reproductive performance during the test period (Table 2). Percent sows bred within 7 days was numerically increased 2.7% in sows in the +Cr group (P =.09) over Control sows. Further, pigs born alive was improved by 0.37 pigs per litter (P =.02) in the +Cr group compared to Controls, and pigs weaned per litter was increased by 0.33 (P >=.02) in the +Cr group. Sows in the +Cr group had a 1.57% lower (P =.11) mortality than Controls. While there were also numerical improvements of varying magnitude in wean-to-first-service interval, nonproductive sow days, and PWMSY for sows in the +Cr group compared to Controls, none of these differences was statistically significant.
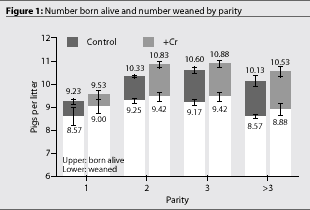
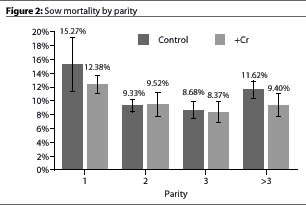
There was no interaction between treatment and parity (P >.48) (Figure 1 and Figure 2).
{kind=link}
{kind=link}
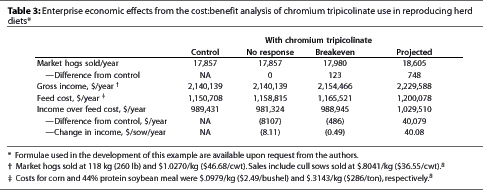
Cost:benefit evaluation
In the "no response" scenario, the total cost of supplementation was $8107 per year or $8.11 per sow per year (Table 3).
{kind=link}
The "Breakeven" scenario demonstrates that an increase of 0.06 pigs weaned per litter resulted in 123 more pigs marketed per year and increased gross income by an amount approximately equal to the cost of chromium supplementation; increasing the response to 0.07 pigs weaned per litter resulted in an increase in income > $0.50 per sow per year.
The "Projected" scenario--which used the performance changes we observed for litter size, wean-to-first-service interval, and sow mortality in the +Cr group--resulted in a mean of 748 more pigs marketed and $40.08 more gross income over feed cost per sow per year for the units included in this study.
Discussion
In the present study, about 100,000 litters were born during the test period. This massive number of litters was reduced to 12 observations in the data set (one observation per unit for the entire year); each data point, then, represents over 8000 litters (100,000 litters on the study / 12 sow units). Averaging so many data points to get one observation per unit tends to minimize the effect of nontreatment factors that normally can have a large impact on interpretation of results. For this reason, the limited PRRSV problems during the test period failed to affect the analysis and the mean response values reported in Table 2 were not adjusted.
The lack of dietary treatment effect during the 6-month loading period was expected. Producers should not expect to see an earlier response in performance records because the mechanism of action that provides for an increase in prolificacy would logically affect the time frame required to see a response. For example, mechanisms of action that affect stillborn numbers might be reflected in a rapid change in production records because it is a near-parturition phenomenon, while mechanisms of action that affected ovulation rate might take >=115 days (an entire gestation period) to produce a change in production records.
The mechanism of action related to CrP is not clearly understood at present (and in all likelihood is not a unidimensional mechanism). Research with CrP conducted in Brazil9 demonstrated a significant increase in viable embryos (litter size at a midgestational slaughter) when the diet of sows was supplemented with CrP at 200 ppb. But this phenomenon seemed to be due to a combination of improved ovulation rate coupled with improved embryo survival, because neither of these parameters, although numerically increased by Cr supplementation, was statistically significant in itself. These findings are consistent with our observation that the significant (P =.02) increase of 0.37 liveborn pigs/litter in the +Cr sows was probably the cumulative result of an increase in total born (0.22), and a decrease in stillborn (.05) and mummies (.10). The numerical decrease in stillborn and mummified pigs is presumably related to late-term uterine conditions that affect fetal survival while the increase in total born pigs is probably also a function of ovulation rate. As with the Brazilian research,9 the P values we calculated for all of the components were not all of the same magnitude. Other studies have demonstrated clear time course effects on the magnitude of response, with greater responses being observed after greater periods of supplementation.2
The improvement in rebreeding values (percent sows bred within 7 days) we observed is consistent with studies from Australia,3,4 and the improvement in litter size (born alive and weaned per litter) is consistent with other studies performed in the United States.1,2 In herds practicing early weaning, in which the average number of litters per sow per year can be expected to be 2.0-2.4, these improvements should increase sow productivity by 0.7-0.9 pigs per sow per year.
The numerical (though not statistical) increase in PWMSY observed was due to the effects on litter size. Pigs weaned per mated sow per year can also be affected by farrowing rate changes, although we did not observe any effect of CrP supplementation on farrowing rate in this study. The lack of a significant difference in farrowing rate between treatments may seem inconsistent with our observation that the number of sows bred by 7 days postweaning was significantly improved in the +Cr group (P =.09). The sows-bred-by-7-days parameter does not evaluate total percentage of sows rebred. The failure to alter farrowing rate in this study accounts for the lack of an observed effect of CrP on nonproductive days.
Other researchers3 have reported that supplementation with CrP reduced death loss from 5% in unsupplemented sows to 1.3% in supplemented sows, while our observations noted a trend toward lower mortality in the +Cr sows. This work in swine is consistent with work in the poultry industry that observed improvements in broiler mortality with supplementation of CrP.10,11 While the mechanism of this action is not totally understood, it may be mediated via the effects of Cr on cholesterol and steroidogenesis. The interrelationship of Cr with cholesterol (from which steroid hormones such as cortisol are built) should not be overlooked. Demonstrations of the effect of Cr on cholesterol and steroids have been observed in multiple species.12-17
The normal effects of parity on litter size are evident in our study. The benefits of the supplementation are demonstrated to be very uniform across the parities for the litter size responses. Although we observed no parity x treatment interactions (P >.33), the effects we found of CrP supplementation on sow measures seem to be dependent on the base performance (Figure 2), with greater responses in situations of lower performance. If this is in fact true biologically, it simply suggests that Cr status is not the sole factor affecting these responses but that it is a contributing factor. It should also be remembered that females at all parities had been supplemented with CrP for a period of time prior to the collection of this data. For parity-one females, supplementation was begun at about 68 kg (150 lb), and for advanced-parity females it was initiated at least 6 months prior to the observations we made in this study.
The mechanism of action of Cr to elicit these metabolic responses has been clarified during the past decade. Evans and Bowman18 have demonstrated an increase in amino acid and glucose uptake by rat skeletal muscle cells due to preculturing of the cells with Cr from CrP, which facilitates more efficient utilization of these nutrients. More recent work19,20 suggests that Cr is involved with insulin via a mechanism that activates a membrane enzyme by a Crbinding protein of low molecular weight that is released concomitant with the insulin response to a meal and also by stimulating insulin-receptor enzyme activity specifically in the presence of released insulin. Garcia, et al.,17 recently observed Cr dose-related effects on tissue sensitivity to insulin in pregnant gilts and found that oxytocin, as well as progesterone (a steroid), were affected by CrP.
Implications
- This study, performed under commercial conditions, demonstrated that supplementing feed with CrP improves litter size and rebreeding efficiency.
- These improvements are observed after continuous feeding during a "loading" period, which was 6 months in reproducing females and from at least 68 kg [150 lb] in replacement gilts in this study.
Acknowledgments
Appreciation is expressed to Steve Abrams for production record compilation and interpretation.
References
1. Lindemann MD, Wood CM, Harper AF, Kornegay ET, Anderson RA. Dietary chromium picolinate additions improve gain/feed and carcass characteristics in growing/finishing pigs and increase litter size in reproducing sows. J Anim Sci. 1995a;73:457-465.
2. Lindemann MD, Harper AF, Kornegay ET. Further assessments of the effects of supplementation of chromium from chromium picolinate on fecundity in swine. J Anim Sci. 1995b;73 (Suppl. 1):185.
5. Steel RGD, Torrie JH. Principles and Procedures of Statistics: A Biometric Approach. 2nd Ed. McGraw-Hill Publishing Co., New York. 1980.
6. Anscombe F, Tukey JW. THe examination and analysis of residuals. Technometrics. 1963;5:141-160.
7. Grubbs FE, Beck G. Extension of sample sizes and percentage points for significance tests of outlying observations. Technometrics. 1972;14:847-854.
8. USDA - NASS. ERS. Agricultural Statistics. 1989 -1998 Annual Summary of Agricultural Prices. 1998.
10. Kim YH, Han IK, Choi YJ, Shin IS, Chae BJ, Kang TH. Effects of dietary levels of chromium picolinate on growth performance, carcass quality and serum traits in broiler chicks. Asian J Anim Sci. 1996a;9:341.
11. Kim YH, Han IK, Shin IS, Chae BJ, Kang TH. Effect of dietary excessive chromium picolinate on growth performance, nutrient utilizability and serum traits in broiler chicks. Asian J Anim Sci. 1996b;9:349.
12. Evans GW, Swenson G, Walters K. Chromium picolinate decreases calcium excretion and increases dehydroepiandrosterone (DHEA) in post menopausal women. FASEB J. 1995;9:525.
13. Page TG, Southern LL, Ward TL, Thompson DL. Effect of chromium picolinate on growth and serum and carcass traits of growing-finishing pigs. J Anim Sci. 1993;71:656-662.
14. Anderson RA. Recent advances in the clinical and biochemical effects of chromium deficiency. In:Prasad AS (ed). Essential and Toxic Trace Elements in Human Health and Disease. Wiley Liss Inc., New York, New York. 1993;221-234.
15. Anderson RA. Chromium. In: Mertz W (ed). Trace Elements in Human and Animal Nutrition (5th ed). Academic Press, Inc., San Diego, California. 1987;225-244.
16. Evans GW. The effect of chromium picolinate on insulin controlled parameters in humans. Int J Biosoc Med Res. 1989;11:163-180
17. Garcia MR, Newcomb MD, Trout WE. Effects of Dietary chromium picolinate supplementation on glucose tolerance and ovarian and uterine function in gilts. J Anim Sci. 1997;75:82(Abstr).
18. Evans GW, Bowman TD. Chromium picolinate increases membrane fluidity and rate of insulin internalization. J Inorg Biochem. 1992;46:243-250.
19. Davis C, Michele K, Sumrall H, Vincent JB. A biologically active form of chromium may activate a membrane phosphotyrosine phosphatase (PTP). Biochem. 1996;35:12963-12969.
20. Davis, C, Michele K, Vincent JB. Chromium oligopeptide activates insulin receptor tyrosine kinase activity. Biochem. 1997;36:4382-4385.
References - nonrefereed
3. Campbell RG. Proc. 16th Annual Prince Feed Ingredient Conf. Quincy, Illinois. 1996;33-38
4. Campbell RG, Purser KW. Proc. 17th Annual Prince Feed Ingredient Conf. Cedar Rapids, Iowa. 1997;15-22
9. Bortolozzo FP, Pinheiro Machado I, Wentz I, Nagae R, Penz Jr AM. Proc. 15th IPVS Congress, Birmingham, England. 1998;79