Original research |
Peer reviewed |
Derivation of porcine circovirus type 2-negative pigs from positive breeding herds
Tanja Opriessnig, DVM; Shan Yu, DVM; Eileen L. Thacker, DVM, PhD; Patrick G. Halbur, DVM, PhD
TO, PGH: Department of Veterinary Diagnostic and Production Animal Medicine, College of Veterinary Medicine, Iowa State University, Ames, Iowa. SY, ELT: Department of Veterinary Microbiology and Preventive Medicine, College of Veterinary Medicine, Iowa State University, Ames, Iowa. Corresponding author: Dr P. G. Halbur, Department of Veterinary Diagnostic and Production Animal Medicine, College of Veterinary Medicine, Iowa State University, Ames, IA 50011; Tel: 515-294-1137; Fax: 515-294-3564; E-mail: pghalbur@iastate.edu
Cite as: Opriessnig T, Yu S, Thacker EL, et al. Derivation of porcine circovirus type 2-negative pigs from positive breeding herds. J Swine Health Prod. 2004;12(4):186-191.
Also available as a PDF.
Summary
Objective: To summarize procedures utilized to derive piglets negative both for porcine circovirus type 2 (PCV2) and for antibody to PCV2, from PCV2-positive breeding herds.
Methods: Sow herds were screened for antibodies to PCV2 by ELISA, with sample-to-positive (S:P) ratios >= 0.2 considered positive. Piglets were obtained by segregated early weaning (SEW) at 10 to 12 days of age. Maternal anti-PCV2 antibody was determined by testing weekly blood samples by ELISA and indirect immunofluorescence assay. Absence of PCV2 was determined by polymerase chain reaction (PCR) on weekly serum samples and by PCR and immunohistochemistry assays on lymphoid tissues collected at necropsy (8 to 15 weeks of age).
Results: Piglets negative for PCV2 by PCR and for anti-PCV2 antibody were derived from PCV2-positive breeding herds. Mean PCV2-antibody half-life in weanlings was estimated to be 19.0 days (95% confidence intervals, 17.6 and 20.3 days). Passively acquired PCV2-antibody present at 10 to 12 days of age declined to S:P ratios < 0.2 by 4.9 +/- 1.2 weeks of age in piglets with S:P ratios >= 0.2 and < 0.6; by 8.1 +/- 1.9 weeks in piglets with S:P ratios >= 0.6 and <= 1.0; and by 11.1 +/- 2.5 weeks in piglets with S:P ratios >= 1.0. Pigs kept in isolation remained PCV2-negative by PCR and ELISA through 8 to 15 weeks of age.
Implications: SEW may be an effective technique to derive PCV2-negative piglets from positive breeding herds.
Keywords: swine, porcine
circovirus type 2, passive antibody decay, ELISA,
segregated early weaning
Search the AASV web site
for pages with similar keywords.
Received: July
3, 2003
Accepted: December
16, 2003
Postweaning multisystemic wasting syndrome (PMWS) is a disease complex characterized by wasting and pneumonia in pigs 5 to 12 weeks old.1 The hallmark lesions of PMWS are lymphoid depletion and histiocytic replacement of follicles associated with porcine circovirus type 2 (PCV2) antigen.2 Transmission of PCV2 is thought to occur through direct contact via oronasal, fecal, and urinary routes.3,4 Vertical transmission has been demonstrated in individual sows in the field5,6 and experimentally.7 There is experimental3 and field8,9 evidence that PCV2 may establish persistent infection in pigs.
Serological surveys in England10 and North America11 found that nearly all swine herds are infected with PCV2. However, detection of antibodies to PCV2 is not necessarily indicative of disease prevalence. Diseases associated with PCV2, such as PMWS, are seen in a relatively low percentage of herds infected with PCV2.1 It has been reported that pigs with sample-to-positive (S:P) ratios of at least 0.6 in an ELISA, using as antigen an open reading frame 2 (ORF2) PCV2 capsid protein, are protected against infection by PCV2 and development of PMWS.12,13
The study described here indicates that, under research conditions, pigs negative both for PCV2 and antibodies against ORF2 of the virus can be derived from PCV2-positive breeding herds. This information may be extrapolated to the field for purposes of establishment of PCV2-free herds.
Materials and methods
Screening of breeding herds for PCV2 by ORF2 ELISA
Screening of blood samples for PCV2-specific antibodies was performed using an ELISA based on a recombinant ORF2 capsid protein as previously described.13 An S:P ratio of >= 0.2 was considered positive (sensitivity = 78.2%; specificity = 99.6%), and an S:P ratio of < 0.12 was considered negative (sensitivity = 90.7%; specificity = 93.2%).13 Values between these S:P ratios were considered suspect for PCV2 antibody. For simplification, the 0.2 cutoff was used to separate seronegative and seropositive piglets in this study.
Six US breeding herds, designated Herds A to F, were screened by testing the dams (sows and gilts) from which pigs were derived approximately 7 to 10 days prior to farrowing, except in the case of Herd B. In Herd B, only gilts were tested, and samples were collected 5 months prior to farrowing. None of the herds tested had a history of PMWS or PCV2-associated disease. The number of sows on each farm randomly selected for screening ranged from 12 to 43, depending on how many piglets were needed for our experiments and how many sows were in the batch ready to farrow at the time piglets were needed.
Selection of dams for derivation of piglets
Where parity information on the dams was available (Herds A, C, D, E, and F), ORF2 ELISA S:P ratios of the dams were compared by parity. Sows within herds were divided into three categories on the basis of prefar-rowing ORF2 ELISA results. Category 1 included negative sows (S:P ratio < 0.2), Category 2 included sows with S:P ratios >= 0.2 and < 0.6, and Category 3 included sows with S:P ratios >= 0.6. Dams were selected for derivation of piglets from either Category 1 or 2. No cross-fostering of piglets was allowed.
Segregated early weaning
The selected piglets were weaned at 10 to 12 days of age and moved to the Livestock Infectious Disease Isolation Facility at Iowa State University (Ames, Iowa) where they were housed. On the day of weaning, blood samples were collected from the anterior vena cava, using a single-use blood collection system (Vacutainer; Becton Dickinson, Franklin Lakes, New Jersey), and tested by ORF2 ELISA. Piglets were randomly assigned to groups and rooms independent of ORF2 ELISA S:P ratios. Handling and housing was the same for all piglet groups. All people entering rooms were required to shower in, put on laundered coveralls, and wear latex gloves when touching piglets. The piglets were kept on raised wire decks, with feed and drinking water provided ad libitum. Airflow was unidirectional (positive pressure), with 10 to 15 air changes per hour.
Monitoring PCV2 status through 15 weeks of age
Randomly selected ELISA-negative samples from 10- to 12-day-old piglets were tested by indirect fluorescent antibody (IFA) assay14 to confirm results. Sera were tested at weekly intervals for PCV2-specific nucleic acids by regular polymerase chain reaction (PCR) and quantitative real-time PCR, using different primer annealing sites as described previously.15,16 At the time of necropsy, performed when pigs were 8 to 15 weeks of age depending on the experiment, lymphoid tissues (lymph nodes, spleen, tonsil, and thymus) were collected in 10% formalin, routinely processed, and tested for PCV2-antigen by immunohistochemistry (IHC) as previously described.17 In addition, blood was collected for ORF2 ELISA testing, and selected lymph nodes were collected, stored at -80°C, and tested by PCR for PCV2-specific nucleic acids.
PCV2 antibody half-life calculation and monitoring of passive antibody decay
The rate of maternal PCV2 antibody decay was determined as previously described.18 A linear regression curve was fitted to the natural logarithms (ln) of the ORF2 ELISA data of 199 randomly selected piglets. The equation h = - (ln 2) b was used to estimate the antibody half-life, where h is the estimated half-life of antibody and b is the slope of the regression line. Antibody half-life of PCV2 antibodies was determined separately for each piglet, and the mean half-life was used as a point source estimate. The 95% confidence interval was calculated on the mean half-life.
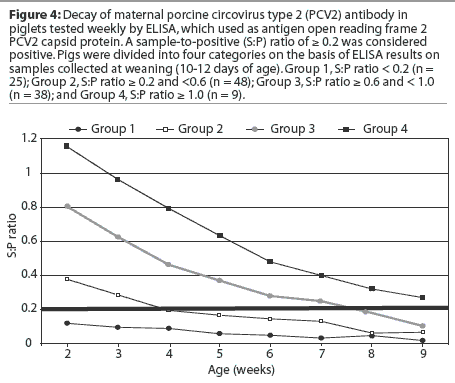
For the purpose of following passive antibody decay, the ELISA S:P ratios of the piglets at 10 to 12 days of age were separated into four categories. Category 1 included seronegative piglets (S:P ratio < 0.2), Category 2 included piglets with low levels of PCV2 antibodies (S:P ratio >= 0.2 and < 0.6), Category 3 included piglets with moderate levels of PCV2 antibodies (S:P ratio >= 0.6 and < 1.0), and Category 4 included piglets with high levels of PCV2 antibodies (S:P ratio >= 1.0). The mean category values were calculated, and passive PCV2-antibody levels were followed over time to determine antibody decay.
Animals and experimental groups
Individual groups of piglets were obtained from Herds A to F for use in several experimental PCV2-inoculation trials performed over a 2-year period. Details of the studies have been described previously.16,19-21 From the time of PCV2 inoculation until necropsy, this report focuses only on the noninfected negative control piglets in these experiments.
Herd A: A total of 98 piglets were obtained from Herd A and brought to the research facility in two batches 3 weeks apart. All piglets were serologically monitored until 6 weeks of age. Negative controls were monitored serologically each week until necropsy: 61 pigs were monitored to 10 weeks of age, 40 pigs to 12 weeks of age, and nine pigs to 15 weeks of age.
Herd B: Seventy-one piglets were obtained from Herd B. All piglets were serologically monitored until 6 weeks of age. Negative controls were monitored serologically each week until necropsy: 28 pigs were monitored to 10 weeks of age, and 14 pigs to 13 weeks of age.
Herd C: Seventy-eight piglets were obtained from Herd C. All piglets were serologically monitored until 8 weeks of age. Negative controls were monitored serologically each week until necropsy at 14 weeks of age (n = 10).
Herd D: One hundred twenty-six piglets were obtained from Herd D. All piglets were serologically monitored until 9 weeks of age. Negative controls were monitored serologically each week until necropsy: 34 pigs were monitored to 9 weeks of age, and 16 pigs to 11 weeks of age.
Herd E: Ninety piglets were obtained from Herd E. All piglets were serologically monitored until 6 weeks of age. Necropsies were not performed on pigs in this group.
Herd F: Forty-one piglets were purchased from Herd F. All piglets were serologically monitored until 5 weeks of age. Negative controls were monitored serologically each week until necropsy: 16 pigs were monitored to 8 weeks of age, and 11 pigs to 11 weeks of age.
Statistical analysis
Summary statistics were calculated to assess the overall quality of the ORF2 ELISA S:P ratios. Welch analysis of variance (ANOVA) was used for parity comparison and for continuous data with unequal standard deviation. The rejection level for the null hypothesis was .05. The R-squared statistic was used to determine what proportion of the total S:P ratio variation of the piglets was explained by the S:P ratio of the dams with S:P ratios < 0.6.
Results
Sow herd PCV2 antibody profiles
Most breeding herds and most sows across parities within herds were seropositive for PCV2 antibodies by ORF2 ELISA (Table 1). There was no difference between high and low parity sows in terms of S:P ratios. The incidence of PCV2 ORF2 antibodies and the within-herd distribution of S:P ratios in the six sow herds are summarized in Figure 1. In the selected herds, 50 to 85% of dams had S:P ratios >= 0.6, 15 to 42% had S:P ratios >= 0.2 and < 0.6, and 0 to 8% were seronegative. Among all breeding herds screened, only one herd (Herd B) was seronegative by the ORF2 ELISA. In this case, all animals tested were gilts, and they were tested prior to breeding and not retested 7 to 10 days before farrowing as for sows from the other five herds.
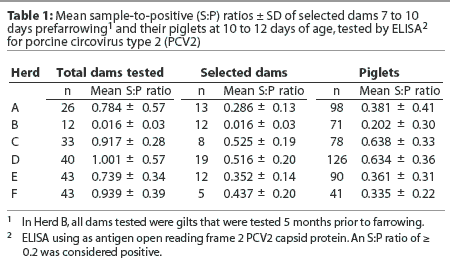
Transfer of passive antibodies from the dams to their offspring
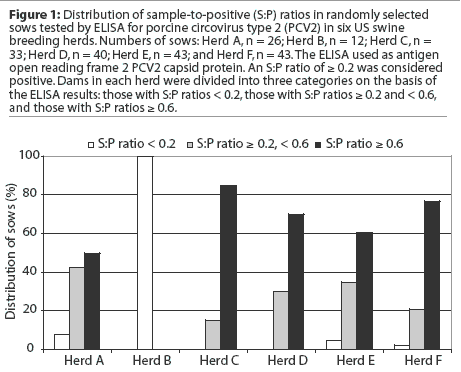
Similar passive antibody transfer data were obtained from all sow herds. Results for sows and piglets are summarized in Table 1. Within the same litter, some individual piglets had up to two to three times higher S:P ratios than their dams, whereas others were seronegative (Figure 2). The r2 for sow S:P ratios and piglet S:P ratios was 0.13, ie, 13% of the variation in the piglets' S:P ratios was explained by the sow S:P ratios (P < .001). Overall, there was limited predictability of passive PCV2 antibody levels in piglets obtained from sows with S:P ratios < 0.6.
Maternal antibody decay in piglets and PCV2 antibody half-life
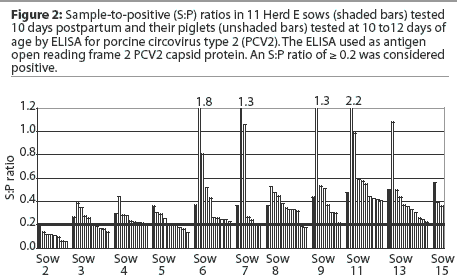
A total of 504 piglets were derived from Herds A to F. Among all piglets, the PCV2 antibody levels at weaning (10 to 12 days of age) were below the ELISA cutoff in 175 piglets (34.7%); within the lower S:P range (>= 0.2 and < 0.6) in 202 piglets (40.1%); moderate (>= 0.6 and < 1.0) in 93 piglets (18.5%); and high (>= 1.0) in 34 piglets (6.7%). The individual distributions of S:P ratios of the piglets in each herd at weaning are summarized in Figure 3.
Within the observation periods, titers of passively acquired PCV2 antibodies declined in all piglets, and none of them seroconverted. The mean antibody half-life, based on declining ELISA S:P ratios in 199 pigs from Herds A to F, was 19.0 days. The 95% confidence intervals for the mean half-life were 17.6 and 20.3 days. In the group with S:P ratios >= 0.2 and < 0.6 at weaning (n = 202), pigs became seronegative at 3 (n = 7), 4 (n = 96), 5 (n = 33), 6 (n = 44), 7 (n = 13), and 8 weeks of age (n = 9), respectively. Piglets with S:P ratios >= 0.6 and < 1.0 (n = 93) became seronegative at 4 (n = 9), 5 (n = 2), 6 (n = 9), 7 (n = 1), 8 (n = 32), 9 (n = 18), 10 (n = 13), and 11 weeks of age (n = 9), respectively. Piglets with S:P ratios >= 1.0 (n = 34) became seronegative at 5 (n = 2), 9 (n = 7), 10 (n = 6), 11 (n = 1), 12 (n = 8), 13 (n = 4), 14 (n = 3), and 15 weeks of age (n = 3), respectively. Antibody levels waned below cutoff levels by 4.9 weeks of age for piglets with low levels of passive antibodies at weaning (95% confidence intervals, 4.8 and 5.1 weeks), at 8.1 weeks of age for piglets with moderate levels of passive antibodies at weaning (95% confidence intervals, 7.7 and 8.5 weeks), and at 11.1 weeks of age for piglets with high levels of passive antibodies at weaning (95% confidence intervals, 10.3 and 12.0 weeks). The antibody decay of 113 pigs is summarized in Figure 4 and represents a typical passive antibody decay curve.
A total of 149 pigs remained segregated and experimentally uninfected, and were necropsied at 8 (n = 5), 9 (n = 18), 10 (n = 35), 11 (n = 27), 12 (n = 31), 13 (n = 14), 14 (n = 10), and 15 (n = 9) weeks of age, respectively. All pigs were free of PCV2 antibody and virus on the basis of results of ELISA and IFA serological testing, PCR on serum and selected lymph nodes, and IHC on lymphoid tissues.
Piglets that were experimentally infected with PCV2 (n = 355) seroconverted within 2 to 4 weeks post challenge and developed PCV2-associated lymphoid depletion and histiocytic replacement of follicles characteristic of PMWS as previously described.16,19-22 PCV2 antigen was detected by IHC in lymphoid tissues associated with lesions, and PCV2-nucleic acids were detected by PCR on serum samples (data not shown).
Discussion
The objective of this report was to summarize the evidence from several experiments that piglets free of PCV2 and antibody to PCV2 can be derived from PCV2-positive breeding herds representative of US swine production. Recently, it has been demonstrated that PCV2-free pigs can be derived by cesarean-section and colostrum deprivation.23 The results of the study summarized here confirm that segregated early weaning (SEW) techniques may also be used for derivation of PCV2-free pigs.
In this study, for the purpose of deriving PCV2-free piglets for research trials, several breeding herds were screened for PCV2 antibodies, and most were positive. Only one breeding herd was seronegative, and these results were based on testing 12 gilts in one farrowing batch.
The use of SEW allowed us to obtain 504 PCV2-free piglets from six seropositive breeding herds with no history of PMWS or PCV2-associated disease. Passive PCV2 antibodies in the piglets, detected by the ORF2 ELISA, decayed to negative levels. The PCV2-negative status of 149 of these pigs was confirmed by PCR on serum and selected lymphoid tissues, as well as by IHC on lymphoid tissues. Most pigs were kept until 10 to 15 weeks of age, and none seroconverted during the observation period.
In contrast to reports of vertical intrauterine transfer of PCV2 resulting in viremic or persistently infected piglets at birth,24 viremia was not detected in these piglets, which were tested weekly by PCR on sera. It has been suggested that piglets can be infected in utero or during the first days after birth.5,6 Vertical PCV2 infection may not always cause fetal death, and virus, antibody, or both were detected in clinically normal piglets.25 It is possible that passively acquired antibody may suppress PCV2 replication until maternal antibody levels wane and the pigs become viremic. The data obtained in our repeated experiments do not support this hypothesis, since all pigs remained seronegative after waning of maternal antibodies.
Transmission routes for PCV2 have not yet been fully investigated. The absence of virus in our pigs may have been due to the carefully controlled environment of the pigs, ie, we can rule out possible transmission by means of semen, equipment, clothing, birds, or rodents in these pigs.
We observed differences in the passive antibody levels within litters. These differences may account for the temporal variability of onset and endemic manifestation of PCV2-associated diseases in many herds. Direct comparison of dam and piglet PCV2 ORF2 ELISA results (Figure 2) suggested that sows with higher S:P ratios tended to have piglets with higher passive antibody levels, and sows with lower S:P ratios tended to have piglets with lower S:P ratios. However, testing was limited to selected dams in the categories with lower S:P ratios. Besides the effect of the sow on the passively transferred antibodies, there might also be a piglet effect, such as failure of colostrum uptake. This might in part explain the lack of maternal antibodies in some piglets derived from seropositive dams (14.1% derived from Herd C, and 8.7% derived from Herd D).
In the United States, clinical PMWS is most often observed in pigs 8 to 12 weeks of age, and, on the basis of experimental models,16,19-22 the pigs are probably infected 7 to 21 days prior to this time. Our results indicate that in piglets weaned at 2 weeks of age, PCV2 maternal antibody decays below ORF2 ELISA cutoff values by approximately 4.9 +/- 1.2 weeks of age in piglets with low levels of antibody at weaning (S:P ratio < 0.6), by approximately 8.1 +/- 1.9 weeks of age in piglets with moderate levels of antibody at weaning (S:P ratio >= 0.6 and < 1.0), and by approximately 11.1 +/- 2.5 weeks of age in piglets with high levels of antibody at weaning (S:P ratio > 1.0).
It has been demonstrated that pigs with ELISA S:P ratio > 0.6 are protected from developing PMWS.12,26 Piglets with lower S:P ratios are therefore at a higher risk for PCV2 infection at an early age. In herds with PMWS, dams with high S:P ratios could be selected to obtain piglets passively protected for a longer time period. However, lack of maternal antibodies is certainly not the only important factor in the pathogenesis of PMWS. Colostrum-deprived, gnotobiotic pigs experimentally inoculated with PCV2 alone did not develop clinical disease.27,28 Other factors, such as coinfections20,29 or certain vaccination protocols,16 may enhance PCV2 infection and must also be considered.
The results of this study may be useful to veterinarians and producers planning to establish a PCV2-free herd. A uniform within-herd distribution of PCV2 antibody in dams might be achieved in the future with use of PCV2 vaccines, resulting in more uniform antibody levels in piglets and appropriate timing of vaccination in growing pigs.
Implications
- Under the conditions of this study, in six breeding herds with no history of PCV2 infection or clinical PMWS, 50% or more of sows randomly tested by serum ELISA had moderate-to-high levels of antibody to PCV2.
- Individual sows with PCV2 ELISA S:P ratios below or close to the positive cutoff (< 0.2) were identified in every herd tested.
- Differences in amount of transfer of maternal anti-PCV2 antibody were observed among piglets within litters.
- Under the conditions of this study, the higher the PCV2 ELISA S:P ratio at weaning, the older the pig when the S:P ratio declined below the positive cutoff value.
- Under the conditions of this study, SEW-derived pigs remained free of PCV2-specific antigen and nucleic acids until at least 15 weeks of age.
- SEW may be an effective technique to derive PCV2-free piglets from positive breeding herds for research or commercial production.
References
1. Harding JCS, Clark EG. Recognizing and diagnosing postweaning multisystemic wasting syndrome (PMWS). Swine Health Prod. 1997;5:201-203.
*2. Sorden SD. Update on porcine circovirus and postweaning multisystemic wasting syndrome (PMWS). Swine Health Prod. 2000;8:133-136.
3. Bolin SR, Stoffregen WC, Nayar GPS, Hamel AL. Postweaning multisystemic wasting syndrome induced after experimental inoculation of cesarean-derived, colostrum-deprived piglets with type 2 porcine circovirus. J Vet Diagn Invest. 2001;13:185-194.
4. Magar R, Larochelle R, Thibault S, Lamontagne L. Experimental transmission of porcine circovirus type 2 (PCV2) in weaned pigs: A sequential study. J Comp Pathol. 2000;123:258-269.
5. O'Conner B, Gauvreau H, West K, Bogdan J, Ayroud M, Clark EG, Konoby C, Allan G, Ellis JA. Multiple porcine circovirus 2-associated abortions and reproductive failure in a multisite swine production unit. Can Vet J. 2001;42:551-553.
*6. Ladekjær-Mikkelsen AS, Nielsen J, Storgaard T, Bøtner A, Allan G, McNeilly F. Transplacental infection with PCV-2 associated with reproductive failure in a gilt. Vet Rec. 2001;148:759-760.
7. Johnson CS, Joo HS, Direksin K, Yoon KJ, Choi YK. Experimental in utero inoculation of late-term swine fetuses with porcine circovirus type 2. J Vet Diagn Invest. 2002;14:507-512.
8. Rodríguez-Arrioja GM, Segalés J, Calsamiglia M, Resendes AR, Balasch M, Plana-Durán J, Casal J, Domingo M. Dynamics of porcine circovirus type 2 infection in a herd of pigs with postweaning multisystemic wasting syndrome. Am J Vet Res. 2002;63:354-357.
9. Larochelle R, Magar R, D'Allaire S. Comparative serologic and virologic study of commercial swine herds with and without postweaning multisystemic wasting syndrome. Can J Vet Res. 2003;67:114-120.
10. Walker IW, Konoby CA, Jewhurst VA, McNair I, McNeilly F, Meehan BM, Cottrell TS, Ellis JA, Allan GM. Development and application of a competitive enzyme-linked immunosorbent assay for the detection of serum antibodies to porcine circovirus. J Vet Diagn Invest. 2000;12:400-405.
11. Magar R, Muller P, Larochelle R. Retrospective serological survey of antibodies to porcine circovirus type 1 and 2. Can J Vet Res. 2000;64:184-186.
*12. Harms PA, Sorden SD, Halbur PG, Nawagitgul P, Lager K, Bolin S, Paul PS. Role of maternal immunity to PCV2 and PRRSV co-infection in the pathogenesis of PMWS. Proc AASV. Kansas City, Missouri. 2002;33:307-311.
13. Nawagitgul P, Harms PA, Morozov I, Thacker BJ, Sorden SD, Lekcharoensuk C, Paul PS. Modified indirect porcine circovirus (PCV) type 2-based and recombinant capsid protein (ORF2)-based enzyme-linked immunosorbent assays for detection of antibodies to PCV. Clin Diagn Lab Immunol. 2002;9:33-40.
14. Allan GM, McNeilly F, Kennedy S, Daft B, Clarke EG, Ellis JA, Haines DM, Meehan BM, Adair BM. Isolation of porcine circovirus-like viruses from pigs with a wasting disease in the USA and Europe. J Vet Diagn Invest. 1998;10:3-10.
15. Fenaux M, Halbur PG, Gill M, Toth TE, Meng XJ. Genetic characterization of type 2 porcine circovirus (PCV-2) from pigs with postweaning multisystemic wasting syndrome in different geographic regions of North America and development of a differential PCR-restriction fragment length polymorphism assay to detect and differentiate between infections with PCV-1 and PCV-2. J Clin Microbiol 2000;38:2494-2503.
16. Opriessnig T, Yu S, Gallup JM, Evans RB, Fenaux M, Pallares F, Thacker EL, Brockus CW, Ackermann MR, Thomas P, Meng XJ, Halbur PG. Effect of vaccination with selective bacterins on conventional pigs infected with type 2 porcine circovirus. Vet Pathol. 2003;40:521-529.
17. Sorden SD, Harms PA, Nawagitgul P, Cavanaugh D, Paul PS. Development of a polyclonal-antibody-based immunohistochemical method for the detection of type 2 porcine circovirus in formalin-fixed, paraffin-embedded tissue. J Vet Diagn Invest. 1999;11:528-530.
18. Paul PS, Mengeling WL, Pirtle EC. Duration and biological half-life of passively acquired colostral antibodies to porcine parvovirus. Am J Vet Res. 1982;43:1376-1379.
19. Fenaux M, Opriessnig T, Halbur PG, Meng XJ. Immunogenicity and pathogenicity of chimeric infectious DNA clones of pathogenic porcine circovirus type 2 (PCV2) and nonpathogenic PCV1 in weanling pigs. J Virol. 2003;77:11232-11243.
20. Opriessnig T, Fenaux M, Yu S, Evans RB, Cavanaugh D, Gallup JM, Pallares FJ, Thacker EL, Lager KM, Meng XJ, Halbur PG. Effect of porcine parvovirus vaccination on the development of PMWS in segregated early weaned pigs coinfected with type 2 porcine cirocvirus and porcine parvovirus. Vet Microbiol. 2004;98:209-220.
*21. Halbur P, Opriessnig T, Hoogland M, Thacker E, Yu S, Meng XJ. PCV2-associated diseases: Research and diagnostic update. Proc Swine Dis Conf Swine Pract. Ames, Iowa. 2003;11:152-160. Available on AASV CD-ROM
22. Fenaux M, Halbur PG, Haqshenas G, Royer R, Thomas P, Nawagitgul P, Gill M, Toth TE, Meng XJ. Cloned genomic DNA of type 2 porcine circovirus is infectious when injected directly into the liver and lymph nodes of pigs: characterization of clinical disease, virus distribution, and pathologic lesions. J Virol. 2002;76:541-551.
23. Tucker AW, McNeilly F, Meehan B, Galbraith D, McArdle PD, Allan G, Patience C. Methods for the exclusion of circoviruses and gammaherpesviruses from pigs. Xenotransplantation. 2003;10:343-348.
24. West KH, Bystrom JM, Wojnarowicz C, Shantz N, Jacobson M, Allan GM, Haines DM, Clark EG, Krakowka S, McNeilly F, Konoby C, Martin K, Ellis JA. Myocarditis and abortion associated with intra-uterine infection of sows with porcine circovirus 2. J Vet Diagn Invest. 1999;11:530-532.
25. Farnham MW, Choi YK, Goyal SM, Joo HS. Isolation and characterization of porcine circovirus type-2 from sera of stillborn fetuses. Can J Vet Res. 2003;67:108-113.
26. Allan GM, McNeilly F, McNair I, Meehan B, Marshall M, Ellis J, Lasagna C, Boriosi G, Krakowka S, Reynaud G, Boeuf-Tedeschi L, Bublot M, Charreyre C. Passive transfer of maternal antibodies to PCV2 protects against development of post-weaning multisystemic wasting syndrome (PMWS): Experimental infections and a field study. Pig J. 2002;50:59-67.
27. Ellis J, Krakowka S, Lairmore M, Haines D, Bratanich A, Clark E, Allan G, Konoby C, Hassard L, Meehan B, Martin K, Harding J, Kennedy S, McNeilly F. Reproduction of lesions of postweaning multisystemic wasting syndrome in gnotobiotic piglets. J Vet Diagn Invest. 1999;11:3-14.
28. Krakowka S, Ellis JA, Meehan B, Kennedy S, McNeilly F, Allan G. Viral wasting syndrome of swine: Experimental reproduction of postweaning multisystemic wasting syndrome in gnotobiotic swine by coinfection with porcine circovirus 2 and porcine parvovirus. Vet Pathol. 2000;37:254-263.
29. Harms PA, Sorden SD, Halbur PG, Bolin SR, Lager KM, Morozov I, Paul PS. Experimental reproduction of severe disease in CD/CD pigs concurrently infected with type 2 porcine circovirus and porcine reproductive and respiratory syndrome virus. Vet Pathol. 2001;38:528-539.
* Non-refereed references