| Original research | Peer reviewed |
Efecto de la estimulación olfatoria durante la lactancia en la conducta agonística en cerdos destetados
SummaryObjective: To determine the effect of olfactory stimulation during suckling on the weaned pig’s welfare. Materials and methods: Six Control group sows and six treatment (Banana) group sows were used, each with 10 piglets. Treatment consisted of daily topical application of isoamyl acetate (banana scent) on the skin of the sows’ mammary glands during the 21 days of suckling. Preference of the piglets for the banana scent was tested with a V-maze at 4 and 21 days of age. After weaning, pigs were regrouped into groups of 10. For the Banana group, feeders were scented with isoamyl acetate for the 15 days after weaning. During the first 5 hours after regrouping, a video camera was used to record nonagonistic, aggressive, and submissive behaviors. Pigs were weighed on their day of birth and at 21 and 35 days of age. Feed consumed was weighed during suckling and for 15 days post weaning. Results: Banana group pigs spent more time in the scented arm of the V-maze. They weighed more after weaning and had greater average daily gain (ADG) than the Control piglets (P < .05). During the postweaning observation period, Banana group pigs showed more nonagonistic and submissive behaviors and less aggressive behavior (P < .05). Feed consumption and ADG did not differ between treatment groups. Implications: Treatment of the sows’ mammary glands with banana scent stimulates milk consumption. Treatment of the feeder with this same scent after weaning is associated with less aggressive behavior, but not with better growth parameters. | ResumenObjetivo: Determinar el efecto de la estimulación olfatoria durante la lactancia en el bienestar del cerdo destetado. Materiales y métodos: Seis hembras del grupo Control y seis hembras del grupo tratamiento (Plátano) fueron utilizadas, cada una con 10 lechones. El tratamiento consistió en la aplicación local diaria de acetato de isoamilo (aroma de plátano) en la piel de la glándula mamaria de las hembras durante los 21 días de lactancia. Se probó la preferencia de los lechones por el aroma de plátano mediante un laberinto en V a los 4 y 21 días de edad. Después del destete, los cerdos fueron reagrupados en grupos de 10. En el grupo Plátano, los comederos fueron tratados con acetato de isoamilo durante los siguientes 15 días después del destete. Durante las primeras 5 horas después de reagrupar a los cerdos, se utilizó una videocámara para registrar la conducta non agonística, agresiva, y sumisa. Los cerdos fueron pesados al nacimiento, el día 21 y 35 de edad. El alimento consumido se peso durante la lactancia y 15 días después del destete. Resultados: Los cerdos del grupo Plátano pasaron más tiempo en el brazo tratado del laberinto. Pesaron más después del destete y tuvieron una ganancia diaria de peso (ADG por sus siglas en inglés) que los cerdos en el grupo Control (P < .05). Durante el periodo de observación, los cerdos del grupo Plátano, mostraron conductas menos agonísticas y sumisa, así como menos agresivas (P < .05). El consumo de alimento y la ADG no difirió entre los tratamientos. Implicaciones: El tratar la glándula mamaria de las hembras con esencia de plátano, estimula el consumo de leche. El tratamiento del comedero con la misma esencia después del destete está asociado con conductas menos agresivas, pero no con mejores parámetros de crecimiento. | ResuméObjectif: Déterminer l’effet de la stimulation olfactive durant la tétée sur le bien-être des porcs sevrés. Matériels et méthodes: Un groupe de six truies témoins et un groupe de six truies traitées (Banane), chacune avec 10 porcelets, ont été utilisés. Le traitement consistant en une application topique quotidienne d’acétate d’isoamyl (arôme de banane) sur la peau des glandes mammaires des truies pendant la période de 21 jours d’allaitement. La préférence des porcelets pour l’arôme de banane a été testée avec un labyrinthe en V alors que les animaux étaient âgés de 4 et 21 jours. Après le sevrage, les porcelets étaient regroupés en groupe de 10. Pour le groupe Banane, les mangeoires étaient aromatisées avec de l’acétate d’isoamyl pour une durée de 15 jours suivant le sevrage. Durant les 5 premières heures après avoir regroupé les animaux, une caméra vidéo a été utilisée pour enregistrer les comportements agonal, agressif, et soumis. Les porcs ont été pesés le jour de leur naissance ainsi qu’à 21 et 35 jours d’âge. La quantité d’aliment consommé était pesée durant la période d’allaitement et pendant les 15 jours suivant le sevrage. Résultats: Les porcs du groupe Banane ont passé plus de temps dans la portion aromatisée du labyrinthe en V. Comparativement aux animaux du groupe témoin ils étaient plus pesants après le sevrage et avaient un meilleur gain moyen quotidien (GMQ) (P < ,05). Durant la période d’observation post-sevrage, les porcs du groupe Banane ont montré plus de comportements non-agonal et de soumission et moins de comportement agressif (P < ,05). La consommation d’aliment et le GMQ ne différaient pas entre les deux groupes. Implications: Le traitement des glandes mammaires des truies avec de l’arôme de banane stimule la consommation de lait. Le traitement des mangeoires avec le même arôme après le sevrage est associé avec moins de comportement agressif, mais non avec de meilleurs paramètres de croissance. |
Keywords: swine, piglet, olfactory stimulation before weaning, isoamyl acetate, behavior after weaning
Search the AASV web site
for pages with similar keywords.
Received: March 18, 2010
Accepted: March 7, 2011
Early weaning (21 days) is very stressful for the piglet and causes an increase in the number of fights among piglets in order to establish social order and hierarchy.1 Fighting may cause the death of some piglets.2 This agonistic postweaning behavior makes the pigs less economically efficient, since feed conversion is poor and growth rate is low. Thus, many researchers have tried to decrease agonistic behavior, for example, through the use of tranquilizers3,4 or pheromones,5,6 design of pens with areas for pigs to hide from and avoid dominant pigs,7,8 regrouping of pigs with different proportions of siblings in each group,9,10 and combination of different litters in the same group during suckling.11-14
Moreover, there has been an attempt to reduce agonistic behavior after weaning by enriching the environment, for example, use of straw bedding, enrichment objects (eg, cotton cords, rubber strips, metal chains), or open-air systems.15-20 These authors reported that environmental enrichment improves social relations and stabilizes the social hierarchy. However, the effect of environmental enrichment by means of early stimulation on the behavior and development of the weaned piglet has never been reported. The method used in this study is early olfactory stimulation of piglets with the intention to create an olfactory attraction conditioned by a given scent (isoamyl acetate) from the moment the piglet is born until post weaning. The hypothesis was that once a positive bond between the suckling piglets and a scent is established, enriching the environment in the nursery pens with this same scent will alter agonistic behavior and development of the pigs when they are regrouped after weaning. The objective of this work was to explore whether learning a neonatal preference during suckling and the presence of this stimulus after weaning have any effect on agonistic behavior and better social integration when the pigs are regrouped after weaning.
Materials and methods
The research protocol was approved by the Bioethical Commission of Murcia University according to the European Council Directives regarding the protection of animals used for experimental purposes.
Animals and facilities
This study was developed in the Teaching Farm of the Veterinary Medicine Faculty, University of Murcia. Twelve multiparous Large White × Landrace crossbred sows were housed in two farrowing rooms with capacity for six animals each (one room per group). Piglets were weighed and identified individually. Litters were balanced, with 10 piglets each, with a minimum weight of 1.20 kg and a similar weight distribution. All sows and litters were housed indoors in farrowing crates (1.7 × 2.5 m total area, with a 0.62 × 2.20-m sow area). Daily management procedures were those generally used in the farms of the area. At 24 hours of age, piglets were routinely processed (ears notched, needle teeth clipped, tails docked, and injected with 200 mg of iron dextran). The farrowing houses were maintained at approximately 20ºC, while the piglets’ area of the farrowing crate was maintained at approximately 30ºC using electric heat lamps. Piglets were weaned at 21 days of age. After weaning, they were housed in rooms each containing six pens (floor area 2.5 m2) with 10 piglets per pen (space allowance 0.25 m2 per pig). The floor was totally slatted and temperature was maintained at 27ºC to 28ºC. During the study, commercial feed was provided, consisting of creep feed from 4 days of age until weaning, and prestarter feed during the 2 first weeks post weaning.
Experimental design and treatments
Study One: preweaning phase. The Control group consisted of six sows (mean parity 3.3, SD 0.5) with 10 piglets each. In order to avoid a differential effect in management (manipulation of the mammary glands and presence of the researchers), the mammary glands of the Control sows were treated with distilled water every day at 9:00 am and
5:00 pm. The Banana group also consisted of six sows (mean parity 3.2, SD 0.4) with 10 piglets each. The two groups were housed in different farrowing rooms. Treatment began on the second day after farrowing and consisted of daily treatment of the skin of the mammary glands at 9:00 am and 5:00 pm with a solution of sunflower oil and isoamyl acetate (banana scent, intended for human consumption), with a ratio of 1:10 sunflower oil to isoamyl acetate. The solution was sprayed on the skin at the rate of 15 mL per sow, measured with a 25-mL graduated cylinder. Control sows were always treated before the Banana group.
Piglets were weighed individually at birth. At 4 days of age, the piglets were again weighed and later were tested in the V-maze, and both groups were provided with plastic creep feeders. In the Banana group, the outside of the feeder was sprayed daily with 15 mL of isoamyl acetate solution. Creep feed was weighed when the feeders were filled, and the remaining feed was weighed at weaning. Piglets were weighed individually at weaning and then were tested a second time in the V-maze.
The V-maze was designed as described by Morrow-Tesch and McGlone,21 Pajor et al,22 and Sevelinges et al.23 The apparatus (Figure 1) included no neutral area: piglets had to choose either the right or left arm of the maze after the start box was opened. The end of one arm contained a grilled window with no olfactory stimulus. The end of the other arm had a similar grilled window where filter paper treated with the isoamyl solution was placed. Filter papers (3 × 3 cm) were saturated with isoamyl solution and stored in a sealed box outside the test room. A fresh filter paper was placed in the arm 15 seconds before each piglet was placed in the start box. The position of the olfactory stimulus (right or left arm) was changed randomly using a table of random numbers (0 = left and 1 = right) generated with the application of Microsoft Excel 2002 (Microsoft Corporation, Redmond, Washington), interchanging the entire ends of both arms and exhausting the inner air space with an electric fan. The upper part of the apparatus was covered with wire mesh to prevent piglets escaping. The floor of the apparatus was a polyvinyl chloride sheet which was replaced for each piglet.
Figure 1: Top view of a V-maze apparatus used to test pigs that had been previously conditioned to a banana scent (isoamyl acetate) and unconditioned pigs (control group). For both groups, a randomly selected filter paper treated with this scent was placed in the grilled section at the end of one maze arm, while a filter paper treated only with sunflower oil was placed in the other arm. The arms and the “choice” area were surrounded by PVC walls 50 cm high. A pig was placed in the start box, and the door was closed behind it once it left the start box. When the four legs of the pig crossed the orange “start-time” line at the threshold of one of the arms, a 3-minute test began, during which the number of times the piglet changed from one arm to the other and the total time spent in the scented arm were recorded. A change of arms was defined as the four legs of the pig crossing the “choice” line (the central orange line in front of the start-box door).  |
The test consisted of placing a piglet in the start box and allowing a few seconds acclimatization time. The door of the start box was opened to allow the piglet to enter the V-maze, and the door was then closed, so that the piglet could choose to go only right or left. The first arm the piglet entered and the time the piglet passed over the start-time line were recorded. A piglet was considered to be inside an arm when all four legs crossed the start line (Figure 1). An observer recorded the pig’s activity, beginning at this point, for 3 minutes using a stop-watch that sounded when the time had passed. During that 3 minutes, the number of times the piglet changed from one arm to the other was recorded, with a change of choice defined as the four legs of the piglet crossing the choice line. The time the piglet remained in the arm with the olfactory stimulus was measured by another stop-watch, and the time spent in the nonscented arm was calculated by subtracting that time from 180 seconds.
The variables analyzed in the V-maze test were the following. Entrance to the V-maze arm was the number of piglets first entering via the scented or nonscented arm. Frequency of arm change was the number of times that a piglet changed from one arm of the V-maze to the other. Time spent in an arm was the total time a piglet spent in that arm.
In order to estimate the possible interference of factors that might influence the results, two preliminary studies with a total of 120 piglets that were not part of the experiment were made before the Banana and Control test in the V-maze. First, the possible preference of pigs for the left or right arm of the maze without any external stimulus was assessed with 30 piglets 4 days of age. Since no differences were found between preferences for the two arms of the maze (P > .05; t-test), it was believed that its construction and layout would not interfere with the results. Second, the possibility was investigated that isoamyl acetate might either attract the piglets or repel them in an innate way, because the piglets used in the preliminary study had never smelled isoamyl acetate. A positive reaction was a neutral reaction, given that neither of these two effects is desired in order to carry out the test. For this test, we used a different group of 30 piglets 4 days of age. It was confirmed that isoamyl acetate neither attracts nor repels piglets that had no previous contact with it (P > .05; t test). Apart from influencing the results, this treatment might also have had a negative effect on the welfare of the piglets, which is contrary to the ethical and legal norms that apply in this kind of experiment. The same results were obtained in preliminary studies conducted in two groups of 30 piglets weaned at 21 days of age.
Study Two: postweaning phase. This study lasted for the first 2 weeks post weaning. In each treatment group, 10 pigs were removed, comprising pigs with diarrhea and pigs with lower body weight. The Control group consisted of the Control pigs from Study One. Control pigs were housed after weaning in five pens of 10 pigs each, with no more than two siblings in each group. The pigs in each pen were each identified with a number (1 to 10) marked on the pig’s back. During the 14 days of the experiment, the pen floor and the external part of the feeder were pressure washed daily with water. Feed was provided ad libitum and weighed whenever feed was added. The Banana group, including 50 pigs from the Banana group in Study One (five pens of 10 pigs each, with no more than two siblings in each group), were placed in a separate room under the same housing and management conditions as the Control group except that the pen floor and feeder were treated with isoamyl acetate solution daily.
For the first 5 hours post weaning, a behavior study was conducted on pigs in the Banana and Control groups. The behavior of the 10 pigs incorporated in every pen of each group was recorded with a camera (Samsung PRO-5780EX/27; Samsung Electronics Co, Ltd) placed 2 m above the floor in order to get an image of the whole pen. In total, 10 different cameras (five for the Banana group and five for the Control group) were connected to a video recorder (Samsung SHR-2162; Samsung Electronics Co, Ltd, Yeongtong-Gu, Suwon City, Gyeonggi-Do, Korea) permitting simultaneous recording of the 10 different cameras. The behaviour study was conducted according to the focal sampling method described by Altmann24 and Peláez del Hierro and Véa Baró.25 The first 10 minutes of every hour recorded by each camera captured the behavior of every pig. Images were studied 10 times for each pen, once for each animal present in the pen. Total time observed for both treatments was 2 groups × 5 pens per group × 10 pigs per pen × 5 first hours × 10 first minutes per hour = 5000 minutes. The ethogram of agonistic behavior applied by Jensen and Wood-Gush26 and later modified by Gonyou27 was used to analyze the pigs’ behavior.
Behavioral measurements
The following nonagonistic behaviors were studied. Feeding: the animal approaches and touches the feeder. It was not possible to distinguish between a productive approach to the feeder (the animal eats some food) and a nonproductive approach (the animal approaches the feeder but eats no food) because of the location of the cameras. Drinking: the animal approaches and touches the water nipple. It was not possible to distinguish between a productive approach (the animal drinks some water) and a nonproductive approach (the animal approaches the water nipple but drinks no water) because of the location of the cameras. Lying: the animal lies laterally, semi-laterally, or abdominally on the floor or on any other animal. Sitting: most of the animal’s weight is on the hindquarters, which rest on the floor. On other animals: the animal passes or stays on top of other animals, either standing or lying. Nose to nose: the nose of a pig approaches the snout or head of the receiver. Nose to body: the nose approaches the body of the receiver. Anal-genital nosing: the nose approaches the anogenital area of the receiver.
The following agonistic behaviors were studied. Six aggressive behaviors were recorded. Fight: rapid and persistent biting in which bites are repeated with a delay of < 1 second between bites, and the aggressive bout lasts not less than 2 seconds. Physical injury is typically inflicted on the recipient. The recipient may or may not respond. Parallel pressing: pressing shoulders against each other, facing in the same direction. Inverse parallel pressing: pressing shoulders against each other, facing in opposite directions. Head-to-head knock: hitting the snout against the head of the receiver. Head-to-body knock: hitting the snout against the body of the receiver. Aiming: an upward-directed thrust of the snout directed at the receiver from a distance of 1 to 2 m.
Five submissive behaviors were recorded. Retreat: takes several steps away from the other animal. Body whirling: the observed individual puts up no resistance to the parallel pressure exerted by the active individual. This behavior is attributed only to the submissive pig. Immobility: the observed individual remains still when it is the object of agonistic actions, with a stance suggesting submission. Hindquarters: the observed animal whirls its body quickly before an attack or a threat, offering the aggressor its hindquarters, with a stance suggesting submission. Head down: the attacked animal puts its head down, an action that is typically submissive.
Statistical analysis
Statistical analyses were performed using the statistical package SPSS for Windows, version 15.0.1, 2006 (SPSS Inc, Chicago Illinois, EE.UU.). For the preweaning phase, a chi-square independence test for two independent samples was performed to compare the proportions of piglets of each group that started the V-maze test choosing the scented or the nonscented arm. The relative risk of entering the scented arm was also calculated. For the variable frequency of arm change, data were transformed by ln(x+1). For the effect of treatment (intergroup differences), a linear mixed model with sow as a random effect was used to analyze intergroup differences in time spent in an arm of the maze, body weight at birth, body weight at 4 days of age, body weight at 21 days of age, average daily gain (ADG), and feed consumption. For differences between Day 4 and Day 21 (intragroup differences), a linear mixed test for related measures was used, with sow as a random effect. Data for all behavioral variables in the postweaning phase were transformed by ln(x+1) and were compared between groups using the linear mixed model with pen as a random effect. The same method was used to analyze the average difference between Control and Banana group piglets in body weight at weaning (21 days of age), body weight at 36 days of age, feed consumption, and feed conversion during the postweaning phase. Values of variables without transformation are presented.
Results
Preweaning phase
Table 1 shows the V-maze test results of the piglets at 4 and 21 days of age. At 4 days of age, the ratio of Control group and Banana group piglets that entered the maze for the first time via the scented arm did not differ. The number of times that piglets changed arms (Figure 2) was significantly less in the Banana group than in the Control group (13.2 ± 3.0 versus 15.4 ± 3.8, respectively; P < .001). In this first test, Banana group piglets spent significantly more time in the scented arm than in the nonscented arm (Table 2). The Banana group piglets also spent significantly more time in the scented arm than did the Control piglets. In the Control group, times spent in the scented and nonscented arms did not differ (P > .05).
Table 1: Number of piglets entering the scented or nonscented arm of a V-maze as a first choice at 4 and 21 days of age*
* Six treatment sows, each with 10 piglets (n = 60 piglets), were treated by daily topical application of isoamyl acetate (banana scent) on the skin of the mammary glands during the 21-day suckling period (Banana Days 0 to 21). Six control sows, each with 10 piglets (n = 60 piglets), were similarly treated with distilled water. The V-maze and testing procedure are described in Figure 1. † Chi-square test of independence for two independent samples was performed to compare the proportions of piglets of each group that chose the scented or the nonscented arm in the V-maze test. ‡ Relative risk and 95% confidence interval (CI) of entering the scented arm of the apparatus as the first choice. |
Figure 2: Frequency of arm change (mean and SD) in the V-maze shown in Figure 1 in tests carried out in piglets 4 and 21 days of age. Six treatment sows, each with 10 piglets, were treated by daily topical application of isoamyl acetate, a banana scent, on the skin of the mammary glands during the 21-day suckling period (Banana Days 0 to 21). Six control sows, each with 10 piglets, were similarly treated with distilled water. A linear mixed model with sow as a random effect was performed to determine intergroup differences (Banana Day 4 versus Control Day 4, P < .01, and Banana Day 21 versus Control Day 21, P < .001). A linear mixed model for related measures with sow as a random effect was performed to determine intragroup differences (Banana Day 4 versus Banana Day 21, P < .001, and Control Day 4 versus Control Day 21, P < .001). 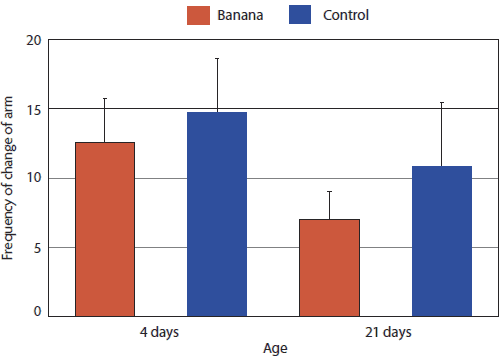 |
Table 2: Total time (mean ± SE) piglets remained in the scented arm of a V-maze apparatus at 4 and 21 days of age*
* Treatments described in Table 1; V-maze described in Figure 1. † Time spent in the maze during the 3 minutes after entry was defined as the total time (seconds) a piglet remained in each arm, with time in the scented arm timed with a stop watch. Time in the nonscented arm = 180 - time in the scented arm (seconds). ‡ Linear mixed model with sow as a random effect. |
||||||||||||||
In the second test, performed at 21 days of age (Table 1), the proportion of pigs that entered the maze via the scented arm was significantly higher in the Banana group than in the Control group (P < .05). The number of times the pigs changed arms (Figure 2) was lower at 21 days of age than at 4 days of age in both groups (P < .001), but Banana group pigs changed arms less often than Control group pigs (6.8 ± 0.2 versus 10.6 ± 0.31, respectively; P < .001). The Banana group pigs spent a longer period of time (Table 2) in the scented arm (P < .001) than in the nonscented arm, while Control group pigs showed a preference for the nonscented arm (P < .001).
The average birth weight of piglets did not differ between the two groups (Table 3). During the period from 1 to 4 days of age, ADG did not differ between groups (Table 3). However, during the period from 5 to 21 days of age, ADG was higher in the Banana group. During this period, the consumption of solid food did not differ between groups (Table 3).
Table 3: Effect of treating the skin of the sow’s mammary glands and the feeder with isoamyl acetate (banana scent) on piglet development and consumption of food during the 21 days of suckling (mean ± SE)*
* Sow treatments described in Table 1. Piglets were weighed at birth, at 4 days of age, and at weaning at 21 days of age. Creep feed was weighed when the feeders were filled at 4 days of age and the remaining feed was weighed at weaning. † Linear mixed model with sow as a random effect. BW = body weight; ADG = average daily gain. |
Postweaning phase
Table 4 shows the main nonagonistic behaviors observed during the first 5 hours after grouping. The frequency of feeding behavior in the Banana group was significantly higher than that in the Control group. Similarly, drinking behavior differed between groups: Banana group pigs made more approaches to the water nipple than did the Control group pigs (Table 4). There was very little feeding or drinking behavior in the Control group during the fifth hour after grouping. The Control pigs lay or sat down more frequently than did the Banana group pigs (Table 4). The maximum difference between groups regarding these two variables occurred during the third hour of observation.
Table 4: Frequencies of nonagonistic behaviors identified in the first 5 hours after weaning in piglets conditioned by treatment of the sow’s mammary glands and the nursery feeder with banana scent*
* Five replicates per treatment (10 pens total) and 10 pigs per pen (n = 50 piglets per treatment). In the Banana group (n = 5 pens), feeders were sprayed with isoamyl acetate (banana scent), once daily during the 14-day experiment. In the Control group (n = 5 pens), feeders were not treated. Banana group pigs had been previously conditioned to the banana scent applied to the skin of the mammary glands of the sow (described in Table 1). Pigs were weaned at 21 days of age. Frequencies (mean ± SE) of nonagonistic behaviors in the observation periods were calculated from observations recorded during the first 10 minutes of each of the first 5 hours after postweaning grouping. The ethogram of agonistic behavior applied by Jensen and Wood-Gush26 and later modified by Gonyou27 was used to record the pigs’ behavior. ab For each behavior, means within a column with different superscripts differ (P < .05). Intergroup differences in each nonagonistic behavior were analyzed using a linear mixed model with pen as a random effect. |
For the variables that showed nonagonistic interactions between pigs, there were no significant differences between the groups, either in the entire 5-hour period or in any sampling period, for the number of times one piglet stayed on top of another. Nose-to-nose, nose-to-body, and anal-genital nosing approaches were considered nonagonistic contacts among the pigs, as no aggressive behavior was observed during or after these contacts. Generally, these behaviors were much more prominent in the Banana group. The sum of these four behaviors (total number of nonagonistic interactions) differed in the second, third, and fifth hours and in the entire 5-hour period.
Table 5 shows agonistic aggressive behaviors. Statistically significant differences in relation to most of the agonistic variables did not occur during the first 3 hours of observation. Within these 3 hours, significant differences between groups were observed only in relation to the head-to-head interaction variable, to the fight variable during the first and third hours, and to the head-body variable during the second hour. Whenever there were differences, the Control group values were always higher than the Banana group values. In the fourth hour, the Control group values for all except the fight variable were higher than those in the Banana group. In the fifth hour, parallel pressings and attacks were more frequent in the Banana group, and head-to-body interactions were higher in the Control group. For the total values for each variable and the hourly total values (Table 5), whenever there were significant differences, values were always higher in the Control group. The variable with the highest values was the head-to-body interactions. For total aggressive behavior, the Control group value was always significantly higher except in the 3rd and 5th hours.
Table 5: Frequency of agonistic aggressive behaviors of pigs detected in the first 5 hours after weaning*
* Treatments described in Table 4. Frequencies (mean ± SE) of aggressive behaviors in the observation periods were calculated from observations recorded during the first 10 minutes of each of the first 5 hours after postweaning grouping. The ethogram of agonistic behavior applied by Jensen and Wood-Gush26 and later modified by Gonyou27 was used to record the piglets’ behavior. ab For each behavior, means within a column with different superscripts differ (P < .05). A linear mixed model with pen as a random effect was used to analyze intergroup differences in each agonistic aggressive behavior. |
Results of the submissive behaviors are shown in Table 6. Three of the five variables (retreat, body whirling, and hindquarters) did not differ between groups. The other two variables (immobility and head down) differed in the first and second hours and overall. In contrast to the aggression variables, when significant differences were found in the variables related to agonistic submissive behavior , the Banana group had the highest values.
Table 6: Frequency of agonistic submissive behaviors of pigs identified in the first 5 hours after weaning*
* Treatments described in Table 4. Frequencies (mean ± SE) of agonistic submissive behaviors in the observation periods were calculated from observations recorded during the first 10 minutes of each of the first 5 hours after postweaning grouping. The ethogram of agonistic behavior applied by Jensen and Wood-Gush26 and later modified by Gonyou27 was used to record the piglets’ behavior. ab For each behavior, means within a column with different superscripts differ (P < .05). A linear mixed model, with pen as a random effect, was used to analyze intergroup differences in each agonistic submissive behavior. |
Table 7 shows growth and consumption of solid food during the first 15 days post weaning. Banana group pigs were heavier than Control pigs both at weaning and at 36 days of age. Growth rate (ADG) did not differ between treatment groups, so that the weight difference at weaning was maintained until the end of the test.
Table 7: Effect of treating the feeder with isoamyl acetate (banana scent) on pig growth and consumption of feed during the 15 days post weaning (mean ± SE)*
* Treatments described in Table 4. Pigs were weaned at 21 days of age. † Intergroup differences analyzed using a linear mixed model with sow as a random effect. BW = body weight; ADG = average daily gain. |
Discussion
At 4 days of age, Banana group piglets did not show an immediate preference for the scented arm of the V-maze, but later recognized and were attracted to the scent, as the total time spent in this arm was higher than that spent in the nonscented arm, in agreement with the work of authors such as Morrow-Tesch and McGlone.21 These investigators found that piglets showed greater preference for an odor they contacted at an early age and were able to identify it during the first few hours after birth. Furthermore, our results suggest that as the piglets matured, the bond between Banana group individuals and the banana odor strengthened. This conclusion is based on the preference of a greater number of Banana group piglets to enter the scented arm first and their significantly lower frequency for changing arms. Furthermore, Banana group piglets spent more time in the scented arm than did Control group piglets. The V-maze test at 21 days of age shows that the Banana group piglets, which had maintained a bond with the banana odor during suckling, opted to remain for a longer period in the scented arm of the maze, which was familiar to them, when they were experiencing a stressful situation (separation from their mother and litter siblings and placement in the V-maze test). These results agree with those of Sevelinges et al23 and Blais et al,28 who tested newborn mice, proving that subjects previously exposed to a specific scent spent more time in the scented arm of a Y-maze labyrinth.
The video-recorded images acquired post weaning showed that the number of visits to the feeder increased with time, in agreement with Campbell,29 who reported that piglets need time to adapt to a new environment and new diet. The number of visits the Banana group made to the feeder was much higher than the number made by the Control group, possibly due to the greater attraction to the feeder treated with the familiar banana scent. Thus, it is possible that the familiar smell favored an earlier and more frequent approach to the feeder in piglets that had adapted to the new environment. However, in this study, it was impossible to discriminate between productive and nonproductive approaches to the feeder because of the placement of the camera. Thus, it cannot be confirmed that the greater number of visits to the feeder by the Banana group meant a greater consumption of feed.
No aggressive behavior was observed when nose-nose, nose-body, or anal-genital nosing interactions were analyzed, as these interactions triggered neither fights nor submissive behaviors. For this reason, these norms of behavior have been included in the nonagonistic category, in contrast to the suggestions of Gonyou.27 In accordance with Morrison et al,30 these behaviors have been considered a type of tactile social interaction or a means of recognition since, as Gonyou31 acknowledges, the sense of smell is the first step in establishing recognition of an individual. In general, in this study, the Banana group showed more exploratory activity in relation to the environment (feeder and drinker) and to their cohorts, while Control group piglets remained in a sitting position or lay down for longer periods of time. According to Fernández32 and Hayne and Gonyou,33 the recumbent or sitting positions are directly related to aggressive agonistic behavior, as confrontations among group members cause physical exhaustion requiring periodic rests for recovery. This aggressive behavior is also in agreement with the results of Arey and Franklin.34 They point out that 40% of the fights observed during the first 5 days post grouping happen within the first 3 hours after grouping has taken place.
The study of agonistic behavior during the first 5 hours post weaning is important because, according to Fenández32 and Hayne and Gonyou,33 the maximum intensity of attacks occurs during this time. Of all the aggressive agonistic behaviors, head-head knocks are remarkable because of the high frequency with which they take place. In this study, the frequencies of head-head knocks were more stable in the Banana group and, with the exception of the fifth hour, when they did not differ, were significantly lower than in the Control group. The phenomenon described by other authors27,35-37 as the existence of the long phases of rest (third and fifth hours) after the intensive fight periods (second and fourth hours), occurred to a greater extent in the Control group. According to Ewbank and Meese38 and Ewbank,39 aggressive interactions taking place within the first hours after grouping are necessary to establish dominance relationships. When dominance relationships have been established, submissive behaviors occur. The results of this study appear to indicate that balance among dominance behaviors and their corresponding submissive behaviors was reached sooner and with a lower aggression level in the Banana group.
The absence of significant differences in birth weight in the two study groups and in feed consumption during suckling seems to indicate that the weight differences obtained in this study in the preweaning phase occurred because Banana group piglets consumed more milk than Control group piglets. Thus, this might support the idea that a positive olfactory determining factor might have been established, as suggested by Sevelinges et al23 in tests carried out on mice. During the first 15 days post weaning, feed consumption, growth rate, and feed conversion were similar in the Banana and Control groups, maintaining the weight difference observed at 21 days of age.
In conclusion, the application of isoamyl acetate to the skin of the mammary glands acted as a stimulus for piglets, favoring a higher consumption of milk. The bond established with the scent on the feeder did not translate into an increase in feed consumption and correspondingly better productive parameters in the postweaning phase. However, the presence of the banana scent during the suckling phase was associated with more stable behavior in these piglets, which can be interpreted as improved welfare by means of a decrease in aggression.
Implications
• Under the conditions of this study, treatment of the skin of sows’ mammary glands with banana scent stimulates milk consumption in the piglets.
• Treatment of the feeder with banana scent after weaning is associated with less aggressive behavior, but not with better growth parameters.
References
1. Olesen LS, Nygaard CM, Friend TH, Bushong D, Knabe DA, Vestergaard KS, Vaughan RK. Effect of partitioning pens on aggressive behaviour of pigs regrouped at weaning. Appl Anim Behav Sci. 1995;46:167–174.
2. Meese GB, Ewbank R. The establishment and nature of the dominance hierarchy in the domesticated pig. Anim Behav. 1973;21:326–334.
3. Gonyou HW, Rohde Parfet KA, Anderson DB, Olson RD. Effects of amperozide and azaperone on aggression and productivity of growing-finishing pigs. J Anim Sci. 1988;66:2856–2864.
4. Tan SSL, Shackleton DM. Effects of mixing unfamiliar individuals and of azaperone on the social behaviour of finishing pigs. Appl Anim Behav Sci. 1990;26:157–168.
5. McGlone JJ, Curtis SE, Banks EM. Evidence for aggression-modulating pheromones in prepuberal pigs. Behav Neural Biol. 1987;47:27–39.
6. McGlone JJ, Morrow JL. Reduction of pig agonistic behaviour by androsterone. J Anim Sci. 1988;66:880–884.
7. Blackshaw JK. The effects of pen design and the tranquilising drug, azaperone, on the growth and behaviour of weaned pigs. Aus Vet J. 1981;57:272–275.
8. Wiegand RM, Gonyou HW, Curtis SE. Pen shape and size: effects on pig behavior and performance. Appl Anim Behav Sci.1994;39:49–61.
9. Friend TH, Knabe DA, Tanksley TD Jr. Behavior and performance of pigs grouped by three different methods at weaning. J Anim Sci. 1983;57:1406–1411.
10. Blackshaw JK, Bodero DAV, Blackshaw AW. The effect of group composition on behaviour and performance of weaned pigs. Appl Anim Behav Sci. 1987;19:73–80.
11. Pitts AD, Weary DM, Pajor EA, Fraser D. Mixing at young ages reduces fighting in unacquainted domestic pigs. Appl Anim Behav Sci. 2000;68:191–197.
12. Weary DM, Pajor EA, Bonenfant M, Ross SK, Fraser D, Kramer DL. Alternative housing for sows and litters: 2. Effects of a communal piglet area on pre- and post-weaning behaviour and performance. Appl Anim Behav Sci. 1999;65:123–135.
13. Weary DM, Pajor EA, Bonenfant M, Fraser D, Kramer DL. Alternative housing for sows and litters: Part 4. Effects of sow-controlled housing combined with a communal piglet area on pre- and post-weaning behaviour and performance. Appl Anim Behav Sci. 2002;76:279–290.
14. Parratt CA, Chapman KJ, Turner C, Jones PH, Mendl MT, Miller BG. The fighting behaviour of piglets mixed before and after weaning in the presence or absence of a sow. Appl Anim Behav Sci. 2006;101:54–67.
15. Blackshaw JK, Thomas FJ, Lee JA. The effect of a fixed or free toy on the growth rate and aggressive behaviour of weaned pigs and the influence of hierarchy on initial investigation of the toys. Appl Anim Behav Sci. 1997;53:203–212.
16. Andersen IL, Andenaes H, Boe KE, Jensen P, Bakken M. The effect of weight asymmetry and resource distribution on aggression in groups of unacquainted pigs. Appl Anim Behav Sci. 2000;68:107–120.
17. Kelly HRC, Bruce JM, English PR, Fowler VR, Edwards SA. Behaviour of 3-week weaned pigs in Straw-Flow, deep straw and flatdeck housing systems. Appl Anim Behav Sci. 2000;68:269–280.
18. Bench CJ, Gonyou HW. Effect of environmental enrichment at two stages of development on belly nosing in piglets weaned at fourteen days. J Anim Sci. 2006;84:3397–3403.
19. Bench CJ, Gonyou HW. Temperature preference in piglets weaned at 12–14 days of age. Can J Anim Sci. 2007;87:299–302.
20. Chaloupkova H, Illmann G, Neuhauserova K, Tomanek M, Valis L. Preweaning housing effects on behaviour and physiological measures in pigs during the suckling and fattening periods. J Anim Sci. 2007;85:1741–1749.
21. Morrow-Tesch J, McGlone JJ. Sources of maternal odours and the development of odour preferences in baby pigs. J Anim Sci. 1990;68:3563–3571.
22. Pajor EA, Rushen J, de Passillé AMB. Dairy cattle’s choice of handling treatments in a Y-maze. Appl Anim Behav Sci. 2003;80:93–107.
23. Sevelinges Y, Lévy F, Mouly A-M, Ferreira G. Rearing with artificially scented mothers attenuates conditioned odor aversion in adulthood but not its amygdala dependency. Behav Brain Res. 2009;198:313–320.
24. Altmann J. Observational study of behaviour: Sampling methods. Behaviour. 1974;49:227–266.
25. Peláez del Hierro F, Veá Baró J. [Ethology: The Biological Basis of Human and Animal Behavior]. Madrid, Spain: Ediciones Pirámide; 1997.
26. Jensen P, Wood-Gush GM. Social interactions in a group of free-ranging sows. Appl Anim Behav Sci. 1984;12:327–337.
27. Gonyou HW. The social behaviour of pigs. In: Keeling LJ, Gonyou HW, eds. Social Behavior in Farm Animals. Ethology: the Biological Basis of Human and Animal Behavior [in Spanish]. Madrid, Spain: Ediciones Pirámide; 1997; Wallingford, UK: CABI Publishing; 2001:147-176
28. Blais I, Terkel J, Goldblatt A. Long-term impact of early olfactory experience on later olfactory conditioning. Dev Psychobiol. 2006;48:501–507.
29. Campbell RG. A note on the use of a feed flavour to stimulate the feed intake of weaner pigs. Anim Prod. 1976;23:417–419.
30. Morrison RS, Hemsworth PH, Cronin GM, Campbell RG. The social and feeding behaviour of growing pigs in deep-litter, large group housing systems. Appl Anim Behav Sci. 2003;82:173–188.
*31. Gonyou HW. Can odours be used to reduce aggression in pigs? Prairie Swine Centre Annual Report. 1997;59–62.
32. Fernández J. [Description of feeding behavior in four pig breeds and study of their relation to productivity, the halothane gene and social hierarchy]. Doctoral thesis. Universitat Autònoma de Barcelona. 2001.
33. Hayne SM, Gonyou HW. Behavioural uniformity or diversity? Effects on behaviour and performance following regrouping in pigs. Appl Anim Behav Sci. 2006;98:28–44.
34. Arey DS, Franklin MF. Effects of straw and unfamiliarity on fighting between newly mixed growing pigs. Appl Anim Behav Sci. 1995;45:23–30.
35. McGlone JJ. A quantitative ethogram of aggressive and submissive behaviors in recently regrouped pigs. J Anim Sci. 1985;61:556–566.
36. Dudink S, Simonse H, Marks I, de Jonge FH, Spruijt BM. Announcing the arrival of enrichment increases play behaviour and reduces weaning-stress-induced behaviours of piglets directly after weaning. Appl Anim Behav Sci. 2006;101:86–101.
37. Colson V, Orgeur P, Courboulay V, Dantec S, Foury A, Mormède P. Grouping piglets by sex at weaning reduces aggressive behaviour. Appl Anim Behav Sci. 2006;97:152–171.
38. Ewbank R, Meese GB. Aggressive behaviour in groups of domesticated pigs on removal and return of individuals. Anim Prod. 1971;13:685–693.
39. Ewbank R. Social hierarchy in suckling and fattening pigs. A review. Livest Prod Sci. 1976;3:363–372.
*Non-refereed reference.