| Original research | Peer reviewed |
Cite as: Smith HM, Selby CC, Williams AM, et al. Effects of day of farrowing induction and spontaneous versus induced farrowing on sow and suckling piglet performance. J Swine Health Prod. 2013;21(4):195–202.
Also available as a PDF.
SummaryObjectives: To evaluate the impact of day of farrowing induction on production parameters under commercial conditions (Analysis 1) and impact of induction, regardless of day, on production parameters (Analysis 2). Materials and methods: First- and second-parity sows (n = 469) were induced with prostaglandin F2α at 3:30 and 6:00 am on gestation day 113, 114, or 116. Sows that had not farrowed by 5:30 am the following day received 10 IU oxytocin. Results: Analysis 1: Gestation length (mean ± standard error) was 114.01 ± 0.08, 114.67 ± 0.08, and 115.19 ± 0.09 days for day 113, 114, and 116 groups, respectively; spontaneous farrowing occurred in 6.50%, 14.89%, and 80.83% of sows. There was no treatment effect on number born alive (NBA), mummies, stillborns, litter weaning weight, litter average daily gain, percentage litters receiving medical intervention once, piglet mortality, or percentage sows in estrus by day 7 post weaning. Percentage unsupervised piglets was greater (P = .001) in day 116 (64.8%) than day 113 sows (56.9%), and intermediate in day 114 sows. Analysis 2: Sows farrowing spontaneously had shorter gestation lengths (114.4 ± 0.09 days) than induced sows (114.7 ± 0.07 days; P = .01). Other sow and piglet parameters did not differ between induced and spontaneous sows. Implications: Under the conditions of this study, more piglets are born during unsupervised hours when sows are induced on day 116 than when sows are induced on day 113. Gestation length must be known for successful farrowing induction. | ResumenObjetivos: Evaluar el impacto del día de la inducción de parto en los parámetros de producción bajo condiciones comerciales (Análisis 1) y el impacto de la inducción, sin considerar el día, en los parámetros de producción (Análisis 2). Materiales y métodos: Se indujeron hembras de primero y segundo parto (n = 469) con prostaglandina F2α a las 3:30 y 6:00 am en el día 113, 114, o 116 de gestación. Las hembras que no habían parido para las 5:30 am del día siguiente recibieron 10 UI de oxitocina. Resultados: Análisis 1: Duración de la gestación (error media ± estándar) fue 114.01 ± 0.08, 114.67 ± 0.08, y 115.19 ± 0.09 días para los grupos del día 113, 114, y 116 respectivamente; el parto espontáneo ocurrió en el 6.50%, 14.89%, y 80.83% de las hembras. No hubo efecto de tratamiento en el número de nacidos vivos (NBA por sus siglas en inglés), momificados, nacidos muertos, peso de destete de la camada, ganancia diaria promedio de la camada, camadas que recibieron intervención médica una vez, mortalidad de lechón, o porcentaje de hembras en estro para el día 7 post destete. El porcentaje no supervisado de lechones fue mayor (P = .001) en las hembras el día 116 (64.8%) que en las del día 113 (56.9%), e intermedio en las hembras del día 114. Análisis 2: Las hembras que parieron espontáneamente tuvieron una duración de gestación más corta (114.4 ± 0.09 días) que las hembras inducidas (114.7 ± 0.07 días; P = .01). Otros parámetros de hembra y lechón no difirieron entre hembras inducidas y de parto espontáneo. Implicaciones: Bajo las condiciones de este estudio, nacen más lechones durante las horas no supervisadas cuando se induce a las hembras en el día 116 que cuando se induce a las hembras en el día 113. Se debe conocer la duración de la gestación para una inducción de parto exitosa. | ResuméObjectifs: Évaluer l’impact du jour d’induction de la parturition sur les paramètres de production sous des conditions d’élevage commercial (Analyse 1) et l’impact de l’induction, indépendamment du jour, sur les paramètres de production (Analyse 2). Matériels et méthodes: Des truies de première et de deuxième parité (n = 469) ont été induites avec de la prostaglandine F2α à 3:30 et 6:00 am au jour de gestation 113, 114, ou 116. Les truies qui n’avaient pas mis-bas à 5:30 am le lendemain matin ont reçu 10 UI d’ocytocine. Résultats: Analyse 1: La durée de la gestation (moyenne ± écart type) était de 114,01 ± 0,08, 114,67 ± 0,08, et 115,19 ± 0,09 jours pour les groupes des jours 113, 114, et 116, respectivement; une parturition spontanée s’est produite chez 6,50%, 14,89%, et 80,83% des truies. Il n’y avait pas d’effet traitement sur le nombre d’animaux nés vivants (NBA), les momies, les mort-nés, le poids de la portée au sevrage, le gain journalier moyen de la portée, le pourcentage de portées ayant reçu une intervention médicale, la mortalité des porcelets, ou le pourcentage de truies en œstrus au jour 7 post-sevrage. Le pourcentage de porcelets non-supervisés étaient plus grand (P = .001) chez les truies induites au jour 116 (64,8%) que celles induites au jour 113 (56,9%), et intermédiaire chez les truies induites au jour 114. Analyse 2: Les truies avec parturition spontanément avaient une durée de gestation plus courte (114,4 ± 0,09 jours) que les truies induites (114,7 ± 0,07 jours; P = .01). Aucune différence n’était notée dans les autres paramètres des truies et des porcelets entre les truies avec parturition induite et celles avec parturition spontanée. Implications: Dans les conditions de la présente étude, plus de porcelets sont nés durant les heures sans supervision lorsque les truies sont induites au jour 116 que lorsque les truies sont induites au jour 113. La durée de la gestation doit être connue pour avoir du succès lors d’induction de la parturition. |
Keywords: swine, farrowing induction, piglet performance, sow performance
Search the AASV web site
for pages with similar keywords.
Received: July 5, 2012
Accepted: December 5, 2012
The use of prostaglandin F2α (PGF) for induction and synchronization of parturition is a common production tool used on sow operations in the United States. Induction facilitates supervision of farrowing by staff, minimizes holiday and weekend work, promotes better cross-fostering, and improves use of farrowing facilities.1-5 Research to develop induction protocols, performed primarily in the 1970s and 1980s, found that the average non-induced gestation length was 114 days. Experiments were conducted comparing sows induced on gestation days 111, 112, and 1135 and 111 and 1126 to non-induced control sows, and no detrimental effects of prostaglandin protocols were found on piglet or sow performance when induction was on day 112 or later. King et al,1 however, reported that piglets born 3 days earlier than non-induced piglets had lower birth weights and 21-day weights. More recently, Gunvaldsen et al,7 inducing on day 114, reported that piglets born on day 115 gained 26 g per day less than piglets born on day 117. They also reported that piglets born on day 115 were 576 g lighter on day 16 of lactation than those born on day 117.7 Other authors have reported an increase not only in lower weights, but also in neonatal loss3 and incidence of splayleg in piglets,8,9 when sows were induced on day 115. Gunvaldsen et al7 reported a tendency toward more preweaning piglet deaths in induced than in non-induced litters. Most piglet deaths occur during the perinatal time frame.10 Spicer et al11 reported that two-thirds of all piglet deaths occur within 4 days of farrowing. It has been reported that the incidence of splayleg increases as gestation length decreases.9 Bölcskei et al8 reported that the number of piglets with splayleg was significantly higher in farrowings induced on day 112 (0.71 ± 1.31) than in those induced on days 113 (0.26 ± 0.53) and 114 (0.18 ± 0.58) and in non-induced farrowings (0.28 ± 0.54). Piglets with splayleg had a higher incidence of death (24%) than did non-splayleg piglets (10%).9 “Normal” gestation length is thought to be longer than 114 days, and in non-induced sows has been reported to be 115-117 days.7,12,13 Inducing on day 112 is likely too early for the modern sow in production systems, causing premature delivery of some piglets. The objective of this study, therefore, was to compare the effects of induction on days 113, 114, and 116 on sow and suckling piglet performance under commercial conditions. A non-induced treatment group was not possible because of potential interruptions of the all-in, all-out flow of farrowing rooms; therefore, day 116 was chosen as the third treatment group, with the expectation that most day 116 sows would farrow spontaneously.
Materials and methods
The standard operating procedures of the study farm were used during the study; therefore, an Animal Care and Use Committee approved protocol was not required.
Animals and facilities
The study was conducted on a new commercial 3000-sow, breed-to-wean farm in central Missouri. All sows were housed in gestation stalls with tunnel ventilation, moved to farrowing rooms with farrowing crates at day 110 of gestation, and had access to water and feed to maintain body-condition score. Post-farrowing feeding was ad libitum. Farrowing rooms were managed all-in, all-out and were ventilated independently using cool cells in the hallway. Sows were arbitrarily allocated to three treatment groups as they entered the farrowing rooms, which consisted of four rows of crates. Each row represented a treatment, and all treatments were represented in each room, with one treatment group represented twice, so that each room had one row for each of two of the treatments and two rows for the third treatment. Inductions started July 14 and ended August 5, 2008.
Study design
Records on 306 first-parity and 163 second-parity sows (469 total) and 5493 piglets born alive were collected. The genetic lines for males and females were G Performer and Genetiporc F20 (Genetiporc, USA, Alexandria, Minnesota), respectively. In Analysis 1, sows were assigned to be induced on gestation day 113, 114, or 116 (Table 1). Parturition induction procedures followed the standard protocol used on the farm prior to this trial, except for day of induction. Briefly, an intramuscular injection of 1.5 mL PGF (Lutalyse; Pharmacia and Upjohn, Kalamazoo, Michigan) was administered at 3:30 am and 6:00 am on day 113, 114, or 116 if sows had not farrowed or showed signs of impending farrowing. Day 116 induction was a proxy for a non-induced group. Day of gestation was calculated from the first day of mating (day 0). As part of the induction protocol, 10 IU of oxytocin was given intramuscularly at 5:30 am the following day to treated sows that had not farrowed.
Table 1: Distribution of first- and second-parity sows induced to farrow on gestation days 113, 114, and 116*
* Data were collected on Genetiporc F20 line sows on a 3000-sow commercial farm in central Missouri, summer 2008. Sows were assigned to treatment (day of farrowing induction) on gestation day 110 when they were moved to the farrowing rooms. A sow that had farrowed or was farrowing on her treatment day was not induced. Sows were induced by intramuscular injection of prostaglandin F2α at 3:30 am and 6:00 am on their assigned treatment day (Analysis 1). |
Farrowing supervision, piglet care, and piglet measurements
Heat lamps and rubber mats were placed in each pen before sows were moved into the farrowing crates. Sows were observed frequently for farrowing and progress from 3:30 am until 2:00 pm. Assistance was provided and oxytocin was administered for obstetrical purposes if progress was not made per the standard operating procedure of the farm. Piglets were cross-fostered within 24 hours to equalize size of piglets and number in litter, within treatment group. At 2 days of age, piglets were weighed and processed. Processing included an iron injection, tail docking, and castration of the males. Litters were observed daily for mortality and morbidity. Dead pigs were removed and the cause of death, determined by farrowing staff, was recorded; however, neither veterinary nor diagnostic testing was done to confirm reason for death. Morbidity was treated on a litter basis, ie, the whole litter was treated if even one pig was in need of treatment. Piglets were weighed again the day before weaning (approximately 21 days of age).
Sow measurements
The farrowing date and time of the first piglet observed relative to the first Lutalyse injection were recorded to determine response. If the first piglet was born prior to the start of the next working day it was assigned the time of ≤ 3:30 am. Birth times were categorized as early (prior to the following working day; < 24 hours), on time (during the following working day; 24 to 35 hours), or late (later than the following working day; > 35 hours). The “on time” category represents the time when sows would likely have farrowed under standard procedures at this farm.
Sows were observed daily for return to estrus after weaning, and percentage returning to estrus within 7 days was calculated. Subsequent reproductive performance was also recorded, though sows not returning to estrus within 21 days of weaning were not included in the analysis for subsequent farrowing. Subsequent farrowing included only sows that farrowed after the first post-weaning mating.
Statistical analysis
Analysis 1. Because the objective was to evaluate what would happen if each of these protocols was applied under commercial conditions, data were analyzed on the basis of the treatment to which sows were assigned, regardless of whether they farrowed spontaneously prior to the day they would otherwise have been induced. An analysis of variance was conducted as a two-by-three factorial (two parities and three treatments) using PROC MIXED in SAS (SAS Inc, Cary, North Carolina) for gestation length, total number born, total number born alive (NBA), mummified fetuses, stillborn, splayleg, prewean death, piglets weaned, litter-processing and weaning weights, litter average daily gain (ADG), and lactation length, with mean differences tested using a Fisher’s least significant difference (LSD) test. The frequency of litter treatments and percentage of sows in heat by day 7 post weaning was analyzed using row-by-column chi-square. The variables odds of being born alive, spontaneous farrowing, unsupervised farrowing, subsequent farrowing, and response to treatment were fitted to a generalized linear model with a logit link function and a binomial distribution, including the effects of treatment and parity as a three-by-two factorial using PROC GENMOD of SAS. Least squares means estimates (average logit) were converted to odds ratios by taking the antilogs of the least squares means estimates. To convert the odds ratio to a percentage, the formula used was (odds ÷ (1 + odds)) × 100. When comparing among treatments, an odds ratio was calculated by taking the antilog of the difference between the least squares means estimates for the two treatments being compared. Converting the data to a logit allowed for the analysis of variance. Significant differences were determined at the P < .05 level.
Analysis 2. Sows were grouped, regardless of day assigned for induction, into those that were induced and those that spontaneously farrowed prior to induction. Sows were retrospectively assigned to treatment groups as having been induced (regardless of day) or having farrowed spontaneously, independent of day of assignment. A mixed-model analysis of variance was conducted as a two-by-two factorial (two parities and two treatments) for gestation length, NBA, mummified fetuses, stillborn, splayleg, prewean death, piglets weaned, litter-processing weight, litter weaning weight, litter ADG, and lactation length, with least squares means differences tested using a Fisher’s LSD. The frequencies of litter treatments and percentage of sows in heat by day 7 post weaning were analyzed using row-by-column chi-square. The variables odds of being born alive and subsequent farrowing were fitted to a generalized linear model with a logit link function and a binomial distribution, including the effects of treatment and parity as a two-by-two factorial.
Results
Analysis 1
Since this was a startup farm, all treatments had more parity-one than parity-two sows (Table 1). Average gestation length was different for all treatment groups (P < .001; Figure 1). The percentage of parity-one and parity-two sows farrowing spontaneously did not differ for induction days 113 and 114 (Figure 2), but in the day 116 treatment, a greater proportion of parity-two sows farrowed spontaneously than did parity-one sows (P < .05). More sows (P < .05) induced on day 113 delivered the first piglet during the following working day than did sows induced on day 114, but neither group differed from day 116 sows (Figure 3). There was also a difference by parity (Figure 4), with a lower (P < .05) percentage of parity-one sows than parity-two sows delivering the first piglet during the following working day.
Figure 1: Gestation length of sows (days from first insemination) by parity and day of induction (Analysis 1). Treatments are described in Table 1. Analysis of variance was conducted as a two-by-three factorial (two parities and three treatments) and mean differences were tested using a Fisher’s least significant differences test. Bars with no common letter differ (P < .001). 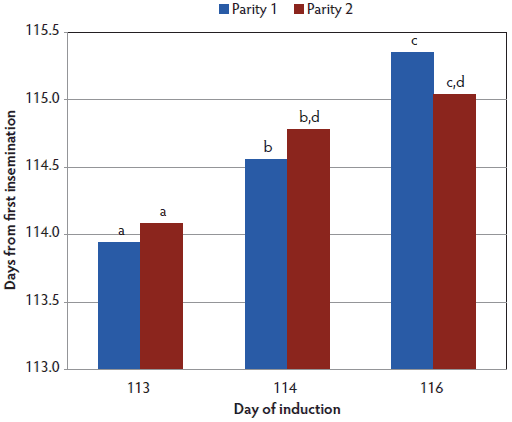 |
Figure 2: Percentage of sows that farrowed spontaneously by parity and day of induction were fitted to a generalized linear model with a logit link function and a binomial distribution including the effects of treatment and parity as a three-by-two factorial. Treatments are described in Table 1. Bars with no common letter differ (P < .05), with significant treatment effect and treatment × parity interaction; a greater proportion of second-parity sows farrowed spontaneously when assigned to be induced on day 116, while the proportions did not differ for first- and second-parity sows assigned to be induced on days 113 and 114. 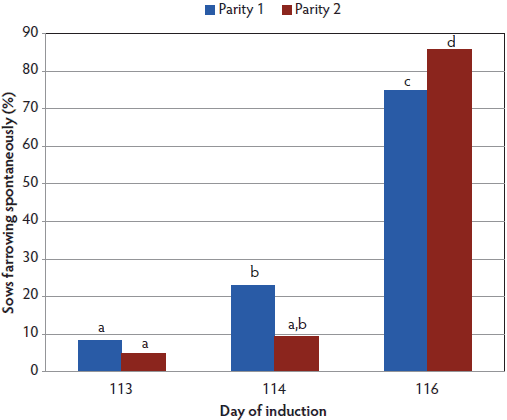 |
Figure 3: Timing of first piglet observed following induction by treatment. Treatments are described in Table 1. Timing of first piglet was fitted to a generalized linear model which included the effect of day of induction in a one-way model with 3 days (SAS PROC GENMOD). Significant differences in timing for proportions of sows farrowing early and on-time (P < .05) are indicated by bars within each timing interval having no common letter. 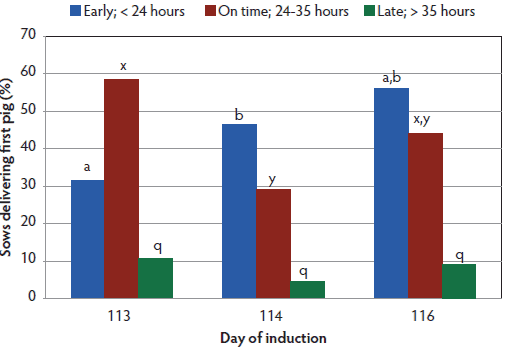 |
Figure 4: Timing of first piglet observed following induction by parity, including litters of 306 first-parity and 163 second-parity sows. Timing of first piglet was fitted to a generalized linear model which included the effect of parity in a one-way model with two levels (SAS PROC GENMOD). Significant differences in timing existed for proportions of sows early and on-time (P < .05) and are indicated by bars within a timing interval having no common letter. Treatments are described in Table 1. 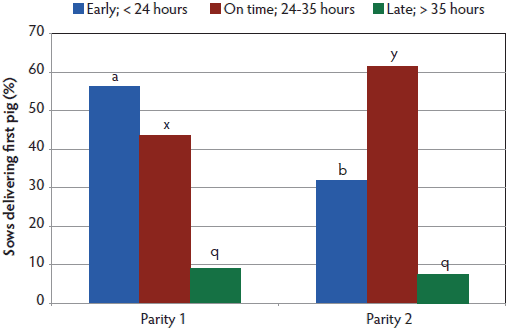 |
There were no significant differences among treatment groups in total number born, NBA, mummies, splayleg piglets, or prewean mortality (Table 2).
Table 2: Least squares means (± SE) of production parameters for farrowing, litter, and sow traits without significant treatment effects (P < .05) (Analysis 1)*
* Treatment and design details described in Table 1. Sows were classified by the day they were assigned to receive an induction protocol (days 113, 114 or 116 of gestation), whether or not they had farrowed prior to treatment. Therefore, a proportion that farrowed spontaneously are included in these means (shown in Figure 2). † Analysis of variance was conducted as a two-by-three factorial (two parities and three treatments) using (PROC MIXED in SAS; SAS Inc, Cary, North Carolina). ‡ Analyzed using row-by-column chi-square (PROC FREQ, SAS; SAS Inc) § Fitted to a generalized linear model which included the effect of day of induction in a one-way model with 3 days (PROC GENMOD, SAS; SAS Inc). ¶ If any pig within the litter was treated, then the entire litter was treated. ADG = average daily gain. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Litters were weighed at processing (day 2) and again the day before weaning, with lactation lengths of 22.10, 21.39, and 21.09 days for days 113, 114, and 116, respectively. Weights at processing differed by treatment (P < .05) and by parity (P = .01). Litters from day 113 sows were lighter at processing than those from day 114 sows, with litters from day 116 sows intermediate (16.20 ± 0.33; 17.36 ± 0.33; 17.08 ± 0.34 kg for day 113, 114, and 116, respectively). Litters from parity-one sows had lighter processing weights than those from parity-two sows, with weights of 16.38 ±.23 and 17.38 ± 0.31 kg, respectively. The number of piglets weaned, litter weaning weights, and litter ADG did not differ (Table 2). The percentages of litters that received medical intervention, either one time or more than once, did not differ among treatments (Table 2). The percentage of piglets that were not supervised was greater (P = .001) for sows induced on day 116 (64.8%) than for sows induced on day 113 (56.9%). Day 114 sows (60.97%) were intermediate and did not differ from the other groups.
Odds of being born alive is equal to the probability of being born alive divided by the probability of being born dead. The analysis showed a numerical treatment × parity interaction, though it did not reach significance (P = .10). This interaction occurred because the odds of being born alive for piglets of the parity-two, day 113 sows were twice that for piglets of the parity-one, day 113 sows. There was no difference between parities in the day 114 and day 116 sows.
The percentage of sows in heat by day 7 post weaning and probability of subsequent farrowing did not differ by treatment (Table 2).
Analysis 2
Induced sows had a longer gestation length (114.7 ± 0.07 days) than did sows that farrowed spontaneously (114.4 ± 0.09 days) (P = .01). Total number born was higher (P < .01) for sows that farrowed spontaneously (13.0 ± 0.30) than for induced sows (12.21 ± 0.26), and sows farrowing spontaneously also tended (P < .10) to have more piglets born alive than induced sows (12.21 ± 0.02 and 11.61 ± 0.18, respectively).
The analysis showed a treatment × parity interaction (P < .05) for litter weights at processing due to litters from parity-two induced sows (17.63 ± 0.37) weighing > 1.5 kg more than litters from parity-one induced sows (16.12 ± 0.29). There was no difference for litters from parity-one and parity-two sows that farrowed spontaneously. Despite differences in lactation length for spontaneously farrowing and induced sows (21.92 ± 0.15 versus 21.31 ± 0.10 days, respectively; P < .001), the number of piglets weaned, litter weaning weights, and ADG did not differ (Table 3). There were also no differences between intervention groups in the incidence of medical intervention one time or more than once, or the odds of being born alive (Table 3). Sow reproductive measures, including the percentage of sows expressing estrus within 7 days of weaning and probability of subsequent farrowing, also did not differ (Table 3).
Table 3: Least squares means (± SE) for production parameters for farrowing, litter, and sow traits with non-significant treatment effects (P < .05) (Analysis 2)*
* Treatment and design details described in Table 1. Sows were classified as having been induced with prostaglandin or having farrowed spontaneously prior to induction. † Analysis of variance was conducted as a two-by-three factorial (two parities and three treatments) using (PROC MIXED in SAS; SAS Inc). ‡ Fitted to a generalized linear model with a logit link function and a binomial distribution, including the effects of treatment (induced versus non-induced) and parity as a two-by-two factorial. § Analyzed using row-by-column chi-square (PROC FREQ, SAS; SAS Inc). ¶ If any pig within the litter required treatment, then the entire litter was treated. ADG = average daily gain.
|
Discussion
The average gestation length for swine has long been accepted to be 114 days, though production data demonstrate a longer average gestation length in today’s sow population, potentially leading to the birth of immature piglets under outdated induction protocols. Parturition induction in swine is accomplished through the use of a luteolytic agent, most commonly PGF or its analog. Prostaglandin F2α has been used to manage reproduction by inducing luteolysis, stimulating contractions of smooth muscle, and stimulating hormonal release from endocrine tissues.4
One of the main reasons for inducing females is to facilitate supervision of farrowing in an attempt to reduce the number of stillbirths.7,14 Cozler et al15 found that the number of litters with stillborn piglets decreased as supervision of parturition increased. When comparing supervised and non-supervised litters, Holyoake et al10 reported that the number of stillbirths per litter was 0.26 for supervised litters and 0.68 for non-supervised litters. In earlier studies,10,16,17 supplemental piglet care was given in addition to obstetrical intervention. In the current study, supervision was given from 3:30 am to 2:00 pm. The higher percentage of supervised piglets for day 113 was expected, because a higher proportion of sows had not farrowed spontaneously and therefore were induced and should have farrowed during supervised hours. It should also be noted that a high proportion of piglets were born during unsupervised hours regardless of treatment, indicating that the farm’s protocol was not successful at meeting the goal of supervision. Although there was a difference in the percentage of supervised piglets, the number of stillborns did not differ among treatments either in Analysis 1 or Analysis 2. Previous research by Ehnvall et al,18 Einarsson et al,19 and Ascher and Tainturier20 has also shown no differences in the number of stillborns between induced and non-induced litters. The level of supplemental piglet care and the proportion of litters actually supervised may explain the differences among studies.
Parity-two sows usually have a longer gestation length than do parity-one sows, which was shown by the relatively higher proportion of parity-two sows being induced versus farrowing spontaneously. Although it is perhaps counterintuitive, spontaneously farrowing sows had a shorter gestation length because they farrowed sufficiently early to preclude induction. The larger litter size may have caused the shorter gestation length of spontaneously farrowing sows,21 and parity-specific protocols may be justified.
It has also been suggested that farrowing induction may decrease preweaning death rate22,23 through better fostering and by ensuring colostrum intake. Hammond and Matty14 reported a reduction of 2.59% in preweaning death rate when sows were induced, compared with the average of the previous 3 years when sows were not induced on the same farms. That study also showed a 1.93% decrease in preweaning death rate compared to that in the previous year.14 Daza et al24 reported a difference in preweaning death rate between parity-two (3.1%) and parity-four sows (2.4%). In the present study, we found no difference in preweaning death rate using either analysis. Consistent with our data, Perez25 found no difference in preweaning death rates among sows induced on day 112 and day 113. Gunvaldsen et al7 also found no difference in the preweaning death rate between induced and non-induced sows.
The odds of being born alive was numerically higher for day-113, parity-two piglets than for day-113, parity-one piglets. The odds of being born alive was not affected by induction in Analysis 2. A better understanding of when to induce by line and parity may improve the value of induction as a tool.
Analysis 1 showed no difference among treatment groups in the total number born, number born alive, or mummified piglets. Alexopoulos et al26 reported a significantly smaller number born alive in litters of sows induced on day 111 than in litters of non-induced sows, but number born alive did not differ between sows induced on day 112 or 113 and non-induced sows. Bölcskei et al8 compared induction later in gestation at days 112, 113, and 114 and found no difference in number born alive among treatments. The effects of induction on number born alive may not have been evident in this study because inductions started later in gestation, on day 113. It is likely that the larger total litter size was associated with the higher frequency of spontaneous farrowing as previously reported.21 We did not expect to see an effect of treatment on the number of mummified piglets, as mummified fetuses occurred before the treatments were applied.
Splayleg, or myofibrillar hypoplasia, is the result of immature development of muscle fibers causing postural collapse of the hind legs and, more rarely, the front legs of afflicted piglets.9 Bölcskei et al8 reported that the occurrence of splayleg was higher in piglets born to sows induced on day 112 (0.71 ± 1.31) than in piglets born to sows induced on day 113 (0.26 ± 0.53) or day 114 (0.18 ± 0.58) or not induced (0.28 ± 0.54). When comparing longer gestation lengths, Walker27 found no noticeable difference in the incidence of splayleg in piglets born to sows with a gestation length of 114.0 versus 116.3 days. The later induction dates in the current study, compared to those of Bölcskei et al,8 might explain differences in results between the two studies.
There has been discussion among producers concerning the vitality and health of induced piglets, with a sentiment that induced piglets have a higher incidence of treatment for sickness. Gunvaldsen et al7 in 2007 showed a greater incidence, almost two-fold, in the percentage of piglets treated. When sows were induced with cloprostenol on day 114, either by intra-abdominal injection (AB) or vulvomucosal injection (VM), 16.1% and 16.3% of piglets were treated, respectively, whereas only 8.3% of control piglets were treated (sows injected AB or VM with sterile saline). Our study showed no treatment effect on the percentage of litters that received treatment once or more than once, although the treatment rate was considerably lower than that of Gunvaldsen et al.7
A measure typically used for reproductive efficiency is the number of pigs weaned per litter. Hammond and Matty14 reported a mean difference of 5.15% extra piglets reared to weaning when sows were induced on days 112, 113, and 114, compared to the average of the previous 3 years (not induced), and 2.68% more piglets compared to the previous year (not induced). Alexopoulos et al26 reported a smaller number of piglets weaned for sows induced on day 111 (8.10 ± 0.27) than for sows induced on days 112 (8.75 ± 0.27) and 113 (9.00 ± 0.16), while Cerne6 reported no difference in the number weaned using the same gestation days. Holyoake et al10 found no difference in litter size weaned between sows induced on day 112 and non-induced sows, nor did Welp et al28 for sows induced on day 111. The current study showed no difference in the number of piglets weaned among treatments for Analyses 1 and 2.
In Analysis 1, litter weights at processing were different across treatment and parity, with no interaction detected. Litters from day 113 sows were lighter at processing than those from day 114 sows, with litters from day 116 sows intermediate. The lack of difference in processing weight in litters from day 116 sows and those from day 113 and day 114 sows might be related to some sows farrowing prior to induction on day 116. Processing weights were lighter in litters from parity-one sows than litters from parity-two sows. In Analysis 2, a treatment × parity interaction for litter weights at processing seems to have resulted because litters from parity-two induced sows weighed > 1.5 kg more than litters from parity-one induced sows, while there was no difference for litters from parity-one and parity-two sows that farrowed spontaneously. There may be a relationship between the growth potential of the piglets, either in utero or after birth, and the milking ability of the sow. A piglet’s growth rate can be used as a measure of vigor and an indication of maturity.29 Jainudeen and Brandenburg12 found that 21-day weights were significantly lower in piglets born to sows induced on days 110 (2.61 ± 0.16 kg) and 112 (2.82 ± 0.36 kg) than in piglets born to sows induced on day 113 (4.26 ± 0.16 kg) or not induced (4.95 ± 0.18 kg). Cerne6 found no differences in weaning weights among piglets of sows induced on days 111 and 112 or non-induced sows. Ascher and Tainturier20 also found no differences among piglets born to sows induced on day 113 with etiproston and cloprostenol and non-induced sows, with weaning weights of 6.9 kg, 6.7 kg, and 6.7 kg, respectively. Our study concurs with those of Cerne6 and Ascher and Tainturier,20 with no differences detected in litter weaning weights. Average daily gain was previously shown to be lower in piglets born to sows induced on days 110 (126 ± 8 g per day) and 112 (130 ± 20 g per day) than in piglets born to sows induced on day 113 (146 ± 8 g per day) or to non-induced sows (143 ± 8 g per day).12 Ascher and Tainturier20 found no differences in ADG when sows were induced on day 113 with either etiproston (218 g per day) or cloprostenol (210 g per day) or farrowed naturally (215 g per day). Again, our data agreed with those of Ascher and Tainturier,20 with no differences among treatment groups for litter ADG in Analyses 1 and 2.
One goal of an induction protocol is to decrease or eliminate variation in timing of farrowing. Jainudeen and Brandenburg12 reported 96% of induced sows delivered their first piglet 28 ± 1.2 hours after the injection. Cerne6 reported > 90% of sows induced on days 111 and 112 delivered the first piglet between 20 and 30 hours later. In the current study, the percentage of sows delivering during the following working day was not as high as previously reported, though the reason for this is not known. Timing of injections should be reconsidered for the current production system.
Consistent with our results, Lebreux et al30 found no difference in wean-to-estrus interval between sows induced on day 113 and non-induced sows, and Cerne6 found no difference in wean-to-estrus interval among sows induced on days 111 and 112 and non-induced sows. The percentages in the present study are relatively low for commercial systems, possibly because the study was carried out in the summer and consisted mostly of parity-one sows. Analysis 2 showed no difference in the numbers of sows displaying estrus by day 7 post weaning. Sows with a wean-to-estrus interval longer than 21 days were not included in the analysis for subsequent farrowing from the first insemination post weaning.
Implications
• Under the conditions of this study, more piglets are born during unsupervised hours when sows are induced on day 116 than when sows are induced on day 113.
• Under the conditions of this study, the day of induction does not affect the number born alive, stillborns, mummies, litter weaning weights, or preweaning mortality.
• Parity-specific farrowing induction protocols merit consideration.
• Under the conditions of this study, sow traits, including the percentage of sows in heat by day 7 and probability of subsequent farrowing, are unaffected by induction treatment.
• It is important to know the average gestation length on a farm prior to initiating a farrowing induction protocol.
• Farrowing induction is a valuable tool when properly applied.
Acknowledgements
The authors acknowledge the cooperation of the owners of the commercial swine facility for the use of their animals.
Conflict of interest
None reported.
References
1. King GJ, Robertson HA, Elliot JI. Induced parturition in swine herds. Can Vet J. 1979;20:157–160.
2. Guthrie HD. Control of time of parturition in pigs. J Reprod Fert. 1985;33:229–244.
3. Dial GD. Clinical applications of prostaglandins in swine. JAVMA. 1984;185:1523–1530.
4. Dial GD, Almond GW, Hilley HD, Repasky RR, Hagan J. Oxytocin precipitation of prostaglandin-induced farrowing in swine: determination of the optimal dose of oxytocin and optimal interval between prostaglandin F2α and oxytocin. Am J Vet Res. 1987;48:966–970.
5. Gall MA, Day BN. Induction of parturition in swine with prostaglandin F2α, estradiol benzoate and oxytocin. Theriogenology. 1987;27:493–505.
6. Cerne F. Induction of farrowing with cloprostenol on a commercial pig breeding farm in Yugoslavia. Vet Rec. 1978;103:469–471. doi: 10.1136/vr.103.21.469.
7. Gunvaldsen RE, Waldner C, Harding JC. Effects of farrowing induction on suckling piglet performance. J Swine Health Prod. 2007;15:84–91.
8. Bölcskei VA, Bilkei G, Biro O, Clavadetscher E, Goos T, Stelzer P, Bilkei H, Wegmüller S. Der Einfluß des Zeitpunktes der Partusinduktion auf das Auftreten der kongenitalen myofibrillären Hypoplasie-Kurzbericht aus der Praxis [The influence of the time of induction of parturition on the appearance of congenital myofibrillar hypoplasia – a short report from practice]. Deutsche Tierarztliche Wochenschrift, 1996;103:21–22.
9. Sellier P, Dando E, Dando P. Induction of parturition in the sow and incidence of splayleg syndrome in the newborn piglet. Annales De Zootechnie. 1999;48:153–161.
10. Holyoake PK, Dial GD, Trigg T, King VL. Reducing pig mortality through supervision during the perinatal period. J Anim Sci. 1995;73:3543–3551.
11. Spicer EM, Driesen SJ, Fahy VA, Horton BJ, Sims LD, Jones RT, Cutler RS, Prime RW. Causes of preweaning mortality on a large intensive piggery. Aust Vet J. 1986;63:71–75.
12. Jainudeen MR, Brandenburg AC. Induction of parturition in crossbred sows with cloprostenol, an analogue of prostaglandin F2α. Anim Repro Sci. 1980;3:161–166.
13. Sasaki Y, Koketsu Y. Variability and repeatability in gestation length related to litter performance in female pigs on commercial farms. Theriogenology. 2007;68:123–127.
14. Hammond D, Matty G. A farrowing management system using cloprostenol to control the time of parturition. Vet Rec. 1980;106:72–75.
15. Cozler YL, Guyomarch C, Pichodo X, Quinio PY, Pellois H. Factors associated with stillborn and mummified piglets in high-prolific sows. Anim Res. 2002;51:261–268.
16. Nguyen K, Cassar G, Friendship RM, Dewey C, Farzan A, Kirkwood RN. Stillbirth and preweaning mortality in litters of sows induced to farrow with supervision compared to litters of naturally farrowing sows with minimal supervision. J Swine Health Prod. 2011;19:214–217.
17. White KR, Anderson DM, Bate LA. Increasing piglet survival through an improved farrowing management protocol. Can J Anim Sci. 1996;76:491–495.
18. Ehnvall R, Einarsson S, Larsson K, Hård af Segerstad C, Westerberg L. Prostaglandin-induced parturition in swine-a field study on its accuracy after treatment with different amounts of PGF2α. Nord Vet Med. 1977;29:376–380.
19. Einarsson S, Fischier M, Karlberg K. Induction of parturition in sows using prostaglandin F2α or the analogue cloprostenol. Nord Vet Med. 1981;33:354–358.
*20. Ascher F, Tainturier D. Etiproston as a part of herd reproduction management. Proc IPVS. Bangkok, Thailand. 1994:376.
21. Rydhmer L, Lundeheim N, Canario L. Genetic correlations between gestation length, piglet survival and early growth. Livest Sci. 2008;115:287–293.
*22. Palomares H, Cervantes A, Lara A, Hernández D. Effects of tiaprost on farrowing performance. Proc IPVS. Lausanne, Switzerland. 1990:449.
23. Stephens S, Boland MP, Roche JF, Reid JFS, Bourke S. Induction of parturition in swine with the prostaglandin analogue fenprostalene. Vet Rec. 1988;122:296–299.
24. Daza A, Bezerra Evangelista JN, Guittiérrez-Barquin M. The effect of maternal and litter factors on piglet mortality rate. Annales De Zootechnie. 1997;48:317–325.
*25. Perez MG. The use of prostaglandins in parturition inductions in the sow. Proc IPVS. Copenhagen, Denmark.1980;51.
26. Alexopoulos C, Saratsis PH, Samouilidis S, Saoulidis K, Kyriakis SC. The effect of cloprostenol alone or with oxytocin on induction of parturition, litter characteristics and subsequent fertility of the sow. Reprod Dom Anim. 1998;33:83–88.
27. Walker N. The effects of induction of parturition in sows using an analogue of prostaglandin F2 α. J Agric Sci Camb. 1977;89:267–271.
28. Welp C, Jöchle W, Holtz W. Induction of parturition in swine with a prostaglandin analog and oxytocin: a trial involving dose of oxytocin and parity. Theriogenology. 1984;22:509–520.
29. Stanton HC, Carroll JK. Potential mechanisms responsible for prenatal and perinatal mortality or low viability of swine. J Anim Sci. 1974;38:1037–1044.
*30. Lebreux B, Ascher F, Tainturier D, Fiéni F. Clinical assessment of a new prostaglandin used for inducing parturition in the sow. Proc IPVS. Bangkok, Thailand. 1994:386.
* Non-refereed references.