Literature review |
January and February, 1997 |
JinLiang Xue, DVM, PhD; Gary D. Dial, DVM, PhD, MBA; James E. Pettigrew, PhD
JLX, GDD: Department of Clinical and Population Sciences, College of Veterinary Medicine; JEP: Department of Animal Science; 385 Animal Science/Veterinary Medicine, 1988 Fitch Ave, University of Minnesota, St. Paul, Minnesota 55108
Citation: Xue JL, Dial GD, Pettigrew JE. Performance, carcass, and meat quality advantages of boars over barrows: A literature review. Swine Health Prod. 1997;1:21-28.
Copyright (C) 1997 American Association of Swine Practitioners.
Also available in PDF format.
Summary
This paper reviews the literature comparing growth performance, carcass, and meat quality data between barrows and boars. Compared with barrows, boars have reduced feed intake, improved feed efficiency, less backfat, higher nitrogen retention, and leaner carcasses. Growth rate of boars will be higher than that of barrows if adequate dietary amino acids are supplied. The improved growth performance and carcass characteristics of boars are primarily due to the anabolic effects of androgens and estrogen produced in the testis. However, administering manufactured anabolic agents, androgens, and estrogens to barrows has not yet been shown to improve growth and carcass in swine.
Keywords: swine, boars, carcass, meat, growth
Received: September 17, 1996
Accepted: December 27,1996
Increased consumer demands for lean pork and heightened awareness of production efficiency have stimulated the pork industry to investigate new strategies to improve lean accretion in pigs being raised to slaughter. Repartitioning agents such as porcine somatotropin or growth hormone (GH), ractopamine, and cimaterol can greatly improve growth rate and feed efficiency, increase lean tissue, and decrease fat deposition.1-3 However, there are some problems with GH. Studies that treated pigs with GH have reported a significant decrease in the tenderness, juiciness, and flavor of pork chops.4-5 Also, pigs treated with GH have increased incidence of pale, soft, and exudative (PSE) pork.6-8 Joint strength and joint soundness is reduced in pigs treated with GH,9 and increased incidence and severity of osteochondrosis has been reported.10 Furthermore, meat tenderness has been reported to decrease in pigs treated with certain b-adrenergic repartioning agents, such as cimaterol.11
Traditionally in most areas of the world, male piglets are castrated shortly after birth despite the fact that intact males are widely reported to have improved feed efficiency and lean accretion over castrated males. Castrating male farm animals is an ancient custom dating back almost as far as human domestication of animals,12 first recorded between the 16th and 11th century BCE in the Chinese Shang Dynasty.13
Several countries, such as Denmark, Britain, Spain, Australia, and New Zealand, currently raise entire males for pork. The European Economic Community (EEC) has passed a regulation (64/433/EEC, 1993) that allows intact male pigs to be reared to 80 kg (176 lb) carcass weight throughout the union countries.14 Carcasses above 80 kg (176 lb) must be tested for odor or taint. In most other countries (including the United States), however, all male pigs not destined for breeding are usually castrated at a young age.
In this review, we discuss the literature comparing growth performance, carcass characteristics, and meat quality of boars and barrows. We also review the other benefits of rearing boars over barrows.
Improved growth performance of intact
male pigs
The relative feed efficiency of boars and barrows reported in the literature has been consistent (Table 1). In general, boars appear to use feed more efficiently than their barrow littermates. Compared with barrows, feed efficiency improvements in boars range from 2.6%-32.1% under various rearing conditions, thus saving 1.8-27 kg (4-60 lb) of feed during the grow-finish period compared to barrows.
The findings on growth rate of boars relative to barrows have been inconsistent. Some researchers found that boars grew faster.15-19 Others did not observe a difference in growth rate between boars and barrows,20-25 and some observed that boars had a lower growth rate than barrows.26-28.
Several factors can influence growth rate in swine and thus should be taken into consideration when the growth rate is compared between boars and barrows. These factors include dietary levels of protein and amino acids, energy intake, age at castration, slaughter weight, and management. The inconsistency of findings in the literature is undoubtedly due at least in part to variability among these factors across studies.
Dietary levels of protein and amino acids
Current literature indicates that boars need more nutrient-dense diets than littermate barrows to achieve improved growth. Current NRC requirements29 are established for barrows and gilts. Increased concentrations of dietary protein over these NRC requirements favor boars, especially at a heavier weight.30 Daily weight gain of boars was reported to be the same as that of barrows on a conventional diet, but boars grew faster than barrows on a higher-protein diet.31 Many experiments have been conducted to determine influences of various dietary protein levels on the growth rate of boars and barrows. The average daily gain (ADG) of boars increased linearly with increasing dietary protein levels ranging from 14%-22% in the growing period up to 55 kg (14 lb) liveweight and from 14%-18% in the finishing period up to 100 kg (220 lb), followed by a decline in gain at 24% or higher protein levels.20,32-35
Lysine is the first limiting amino acid for growing pigs fed corn-soy diets. Boars require a higher level of dietary lysine than barrows.30-32,36 Significant reductions in growth rate, feed efficiency, and nitrogen retention are evident in boars fed low-lysine diets (0.47%-0.60%). When bodyweight is between 25-55 kg (55-124 lb), there is a linear increase in the growth rate and improved feed:gain ratio (F:G) as dietary lysine concentrations increase from 0.75% to 1.11%.21,35,36 The concentration of lysine required to achieve maximum gain in boars is about 11.8 g per kg (25 g per lb) when bodyweight is between 25-55 kg (55-124 lb). Barrows and gilts require lower lysine concentrations when in the same range of bodyweight.37 During the 50-90 kg (110-198 lb) liveweight phase of growth in boars, there is a linear response for the growth rate, F:G, and carcass lean proportions when lysine concentrations are increased up to 9.1 g per kg (20 g per lb) with a daily lysine intake of 22.9 g per kg (50 g per lb) for boars.38 It is important to remember that amino acid requirements vary among boars as well as among barrows due to differences in genetic lines, health status, environment, etc.
An excessive protein intake may also result in depressed boar growth performance.32,35,39,40 Besides the reduced voluntary feed intake, the growth-decreasing effect of high-protein diets is partly due to the reduced energy value of diets with increased protein concentrations.41,42 Excess protein intake causes a high rate of protein turnover, increased muscle respiration43 and increased visceral organ mass,44,45 which contribute to increased heat production.46 It appears that a supply of protein in excess of requirements reduces the net energy available to animals, which in turn depresses the rates of deposition of protein and water, and to a lesser extent, the deposition of fat.35,40 The same effect has been observed in castrated male pigs given high protein diets ad libitum.47
Energy intake
When fed ad libitum, barrows consumed more feed than boars and exhibited a liveweight gain similar to that of boars.48 When energy intake was restricted, boars grew faster than barrows from 45-90 kg (100-200 lb) liveweight.48 It is clear that boars grow faster than barrows when fed restricted diets, and barrows grow faster than boars when fed ad libitum.49,50 The association of growth rate with feeding patterns is probably related to appetite. Several researchers have reported that barrows have greater appetites than boars,15,20,26,50 i.e., they have increased daily feed intake compared to boars when animals are given free access to feed. The difference in appetites is profound from 55 kg (121 lb) liveweight onwards.21 A dramatic increase in backfat thickness of barrows from 56 kg (123 lb) liveweight (120 days old) has been reported, but this increase did not occur in entire males.28 Further research needs to be conducted to determine whether the differences in body composition between boars and barrows are due to feed intake, or vice versa.
Age at castration
Age at castration may affect growth performance of barrows. It seems that superior muscle growth in entire males is manifested only when animals are approaching puberty. Castration did not affect liveweight or total soft tissue protein content when performed at 1, 2, or 4 weeks of age.51 In barrows that had been castrated at 15 and 40 kg (33 and 88 lb) bodyweight, skeletal muscle protein accretion did not differ from that of boars 5 weeks after castration. There was no difference in ADG among barrows castrated at birth or at 6, 12, 16, and 20 weeks of age52 and at 3-4, 9-10, and 15-16 weeks of age.53 Average feed intake of barrows castrated at 16 and 20 weeks of age was decreased compared with those castrated at birth or at 6 and 12 weeks of age.52 In contrast, postpubertal boars had greater muscle protein accretion rates than barrows that were castrated at 75 kg (165 lb) liveweight.54
Slaughter weight
In many European countries, 90-100 kg (198 - 220 lb) liveweight has been regarded as the optimal slaughter weight. In the United States, however, slaughter weights tend to be heavier, and slaughter at 120-130 kg (264-286 lb) is not uncommon. In a Swedish study of two genetic lines (Landrace and Yorkshire),55 boars reached slaughter weights of 90, 110, and 130 kg (198, 242, and 286 lb) earlier than barrows and gilts. The magnitude of the differences in age to attain a given slaughter weight are greater between barrows and boars as bodyweight increases. Boars attained a slaughter weight of 110 kg (242 lb) 10 days sooner than barrows.55 When fed to higher slaughter weights (e.g., 130 kg/286 lb), boars had 10% higher ADG than barrows. Boars had better feed efficiency than barrows from 70-130 kg (154-286 lb) bodyweight. The ADG of barrows reached a maximum from 70 to 90 kg (154 to 198 lb) liveweight, after which their ADG decreased. In boars, ADG peaked at a bodyweight 10 kg (22 lb) heavier than for barrows, and the magnitude of the subsequent decrease in ADG was smaller in boars compared to barrows after that weight.15,55 In a similar study in the United States,24 ADG of barrows was slightly greater than boars until 76 kg (167 lb) liveweight (17 weeks old), when it plateaued. Boars reached the maximum rate of gain at 87 kg (192 lb) (20 weeks old). Thus, the bodyweight at which ADG peaked was 11 kg (24 lb) heavier for boars than for barrows.
Rates of nitrogen deposition also appear to differ between barrows and boars. The rate at which nitrogen was deposited responded quadratically over time for both boars and barrows as they gained weight, reaching a maximum for boars at a liveweight of 60 kg (132 lb) and for barrows at 55 kg (121 lb).56 The difference in weight/age of maximum growth rate and nitrogen deposition between boars and barrows is another explanation for the inconsistencies of growth rate found in the literature.
Management
Floor space and group size affect the growth rate of pigs. The ADG of pigs with a floor space of 0.34 m2 (3.7 ft2) per pig was reduced compared to pigs that had 0.68 or 1.01 m2 (7.3 or 10.9 ft2) per pig.57 Meunier-Salaun, et al.,57 determined the optimal range of floor space to ensure optimal performance. We speculated that pig density may have been a primary factor causing reduced ADG of boars compared to barrows in some studies.28 Aggressive behavior increases when group size and pig density increase.58 Swine with higher concentrations of salivary androstenone are more aggressive than those with lower concentrations.59 Group size (40 pigs per pen)28 may influence growth rate of boars because they have higher concentrations of androstenone than barrows.
No management problems were reported when boars and gilts were raised together up to bodyweights of 81, 93, and 110 kg (178, 205, and 242 lb). At heavier weights, growth performance was slightly reduced for the boar/gilt groups compared to mixed barrows and gilts.60,61
Carcass comparison
Reports in the literature of carcass comparisons between boars and barrows are consistent (Table 2). Boars have less backfat,16,20 longer carcass length,24,62 and a larger longissimus muscle area than do barrows.21,22,30 Barrows tend to have a higher dressing percentage compared with their contemporary intact males, due partly to the higher genitalia weight of boars.20,60-64
Boar carcasses have more lean meat, less fat, more bone, and more skin compared to barrows (Table 3). The carcass composition of gilts is intermediate between boars and barrows. Ratio of bone:muscle is relatively constant at between 1:5.4 and 1:5.6 in boars and barrows.20 Boar carcasses contain more water and protein and less fat than those of barrows (Table 4).
Lean meat
Boars retain a higher percentage of nitrogen intake and of digested nitrogen than barrows when they are given a diet with concentrations of amino acids that support the higher nitrogen retention.36,40 Castration reduced the ceiling for protein deposition in one study by 30%, from 128 to 85 g (4.5 to 3.0 oz) per day.48 Castrated male pigs have an inherently lower capacity for protein deposition and a higher rate of fat accretion, and thus require less dietary protein and amino acids to support maximum growth than entire males.20,34,36-38,65
Proportions of lean meat and adipose tissue in the carcass are affected by the interaction between gender and amino acid levels. The amino acid requirements for lean accretion is higher for boars than for barrow littermates.30,31 Dressing percentage decreases at a similar rate for both boars and barrows as dietary protein increases.63,66,67
With an adequate protein and lysine diet, protein deposition of boars increased linearly with increased energy intake up to 7.9 mcal DE per day, and then remained constant when feed intake was at 120 g per day thereafter. The response in barrows was similar to that of boars, but protein deposition in barrows reached a maximum value of only 85 g per day.48
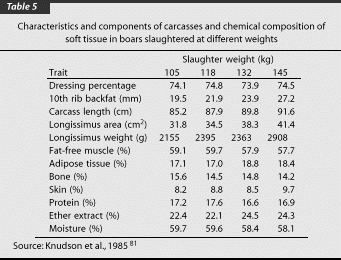
Rearing pigs to heavier slaughter weights results in a lower percentage of lean meat in carcasses. The effect of slaughter weight on lean percentage is, however, not the same in barrows, gilts, and boars. Lean percentage in barrows and gilts is more influenced by weight at slaughter, while in boars slaughter weight is less of a factor in lean percentage.55 At 90 kg (200 lb) liveweight, each Landrace boar produced 3.7 kg (8.1 lb)more lean meat than did a 90-kg (200-lb) barrow. The magnitude of the difference between boars and barrows increased as slaughter weight increased. At 130 kg (286 lb) liveweight, boars had 7 kg (15.4 lb) more lean meat than barrows. With a higher lean meat content in boar carcasses, the content of fat in boars is lower than that in barrows.49,55 For all traits, gilts were intermediate between barrows and boars. As liveweight of boars increased from 105 to 145 kg (230 to 320 lb), carcass length, tenth rib backfat thickness, longissimus muscle area, and longissimus weight increased linearly. Proportion of moisture and protein in the soft tissue decreased while ether extract increased as liveweight increased in boars (Table 5).
{kind=link}
Fat tissue
Boars have been reported to have softer fat than barrows.68,69 Soft fat fails to "set" properly after chilling, as tissue separates between the fat-fat or fat-muscle interfaces.70 Edwards, et al.,71 reported that the gender effects on soft fat are not evident when fat hardness is adjusted for fat thickness. However, Dransfield and Kempster72 observed that boars have softer fat than gilts and barrows even after adjusting for fat thickness. Leaner pigs have softer fat, but castration had no effect on fat hardness in pigs at 68 kg (150 lb).73
Backfat from intact males, compared with barrows and gilts, has a relatively higher proportion of unsaturated fatty acids, mainly linoleic acid.52,74 The highest proportion of saturated acids are observed in barrows, mainly due to a higher palmitic acid content. Gilts are intermediate between barrows and boars.70,75,76 It has been suggested that this difference may result in a softer backfat and marbling of boars. In all sexes, palmitic, stearic, oleic, and linoleic acids are the major fatty acids and amount to over 90% of the total fatty acids in the backfat.77
Bone and skin
Boars have heavier bones than barrows at the same slaughter weights. The difference in bone content between boars and barrows is approximately the same at several slaughter weights.55,78,79 With increasing weight, the percentage of bone decreases. The greater proportion of bone may be due, at least in part, to the anabolic effect of sexual hormones, such as testosterone, on the growth rate of specific bones and body components.
Boars have thicker skin than barrows.17,18,80,81 The dermis above the first lumbar vertebra in boars was 0.82 mm (0.03 inches) thicker than that in barrows (3.76 versus 2.94 mm, 0.15 versus 0.12 inches) at 90 kg (200 lb) liveweight in the Landrace genotype.80 In addition to the difference in thickness, the dermis of barrows has a finer structure than that of boars. Sebaceous glands in barrows are poorly developed.80
Meat quality
Weight loss through exudate or drip is an economically important meat quality trait in pigs.82 Carcass pH values at 45 minutes (pH45) and 24 hours (pHu) post slaughter are widely used as indirect measurements of incidence of PSE and dark, firm, and dry (DFD) meat quality defects. Abnormally low pH45 values are associated with PSE while abnormally high pHu values typically occur in DFD meat. Pork with a pH45 < 6.1 has higher water drip while pork with a pH45 > 6.1 has improved water holding capacity and less water drip.83 Boars have been reported to have higher pHu values than barrows.84 In boars held overnight after transport before slaughter, pH values and DFD frequency were higher compared with barrows.85,86 During preslaughter holding, physical activity is associated mainly with aggressive behavior. Entire males tend to be more aggressive than barrows. There is no difference in DFD and PSE frequencies between boars and barrows when less than 4 hours elapse from farm to slaughter.75
Other advantages of rearing intact males
Genetic selection
Because most male pigs are castrated at young ages, breeding stock must be selected at an early age before their true genetic potential can be accurately assessed and only a limited number of boars are tested for genetic improvement. If boars were reared for meat, genetic companies would have larger populations for genetic selection.
Castration and animal welfare
Raising boars removes labor input for castration. Castration is believed to be stressful and painful to the animals, which raises ethical and welfare concerns.87 Legal restrictions on animal welfare are already in place in some European countries.88
Efficiency of nutrient use
Swine farm waste and related odor are becoming serious issues in the swine industry, particularly in the top pig production states, such as Iowa and North Carolina.89,90 It was recognized that animal production causes almost 90% of the total NH3 emission.91 In Denmark, nitrogen content in manure has been used for determining the amount of manure applied per hectare of land.92 Since boars have improved feed efficiency and nitrogen retention compared with barrows,36,40,56 avoiding castration would improve nitrogen utilization in swine production systems.
Financial return
Packers pay a much lower price for boar carcasses than carcasses from barrows. In recent upper Midwest markets, packers paid approximately $0.22 per kg ($0.10 per lb) of carcass weight less for boars than barrows produced from the same genetic company, indicating that producers will receive $22-$28 less for a boar than a barrow--a considerable economic loss for producers. If boars were sold at the same price as barrows, based on our data,28 each boar would net $5-$20 more than a barrow because of improved feed efficiency and lean proportion in carcasses.
Anabolic effect of gonadal steroids
The superior performance and carcasses of boars relative to barrows are explained by the anabolic effect of gonadal steroids. Anabolic agents used in beef cattle result in increased feed intake, daily weight gain, and carcass lean proportion, and decreased F:G.93,94 However, when administered after castration, manufactured anabolic agents, exogenous androgens or/and estrogens had negligible effects on growth performance in pigs in some studies,95-104 and positive effects on growth performance in other studies.105 The reason for interspecies variation in the effects of anabolic agents is not yet determined.
Implications
- Intact male pigs have improved feed efficiency and a higher proportion of lean tissue than castrated males.
- Intact males grow faster than barrows when diets with adequate concentrations of protein and amino acids are supplied.
- Producers will receive a higher net financial benefit from rearing intact male pigs than that from barrows if boars and barrows were sold at the same price.
References
1. Chung CS, Etherton TD, Wiggins JP. Stimulation of swine growth performance by porcine growth hormone. J Anim Sci. 1985; 60:118-130.
2. Etherton TD, Sorensen MT, Coleman ME, Chaudhuri S. Mechanisms by which somatotropin alters muscle growth and adipose tissue accretion. Proc Recip Meats Conf. 1989;42:83-89.
3. Boyd RD, Bauman DE, Fox DG, Scanes CG. Impact of metabolism modifiers on protein accretion and protein and energy requirements of livestock. J Anim Sci. 1991;69 (Suppl. 2):56-75.
4. Boles JA, Parrish FC, Jr Shaggs CL, Christian LL. Effect of porcine somatotropin, stress susceptibility and final end point of cookery on the sensory, physical, and chemical properties of pork loin chops. J Anim Sci. 1991;69:2865-2870.
5. Boles JA, Parrish FC, Jr Shaggs CL, Christian LL. Sensory, physical, and chemical properties of pork loin chops from somatotropin-treated pigs of three stress classifications. J Anim Sci.1992; 70:3066-3070.
6. Solomon MB, Campbell RG, Steele NC, Capema TJ, McMurtry JP. Effect of feed intake and exogenous porcine somatotropin on longissimus muscle fiber characteristics of pigs weighing 55 kilograms liveweight. J Anim Sci.1988;66:3279-3284.
7. Soloman MB, Campbell RG, Steele NC, Caperna TJ. Effects of exogenous porcine somatotropin administration between 30 and 60 kilograms on longissimus muscle fiber morphology and meat tenderness of pigs growth to 90 kilograms. J Anim Sci.1989;67(Suppl. 1):153(Abstr).
8. Solomon MB, Campbell RG, Steele NC. Effect of sex and exogenous porcine somatotropin on longissimus muscle fiber characteristics of growing pigs. J Anim Sci.1990;68:1176-1181.
9. He P, Aherne FX, Nam DS, Schaefer AL, Thompson JR, Nakano T. Effects of recombinant porcine somatotropin (rpST) on joint cartilage and axial bones in growing and finishing pigs. Can J Anim Sci.1994;74:257-263.
10. He P, Aherne FX, Schaefer AL, Thompson JR, Nakano T, Jones SDM. Differentiation of the effects of somatotropin and enhanced growth rate on the occurrence of osteochondrosis in pigs. Can J Anim Sci.1994.74:251-255.
11. Jones RW, Easter RA, McKeith FK, Dalrymple RH, Maddock HM, Bechtel PJ. Effect of the b-adrenergic agonist cimaterol (CL 263,780) on the growth and carcass characteristics of finishing swine. J Anim Sci.1985;61:905-913.
12. Turton JD. The effect of castration on meat production and quality in cattle, sheep and pigs. Anim Breed Abstr.1962;30:447-456.
13. Zhang ZG, Li BT, Chen XH, et al. Pig Breeds in China. Shanghai Scientific & Technical Publishers, Shanghai, China.1986; p. 1-6.
14. Willeke H. Possibilities of breeding for low 5b-androstenone content in pigs. Pig News Info. 1993;14:31N-33N.
15. Blair R, English PR. The effect of sex on growth and carcass quality in the bacon pig. J Agric Sci Camb.1965;64:169-176.
16. Wong WC, Boylan WJ, Stothers SC. Effects of dietary protein level and sex on swine performance and carcass traits. Can J Anim Sci.1968;48:383-388.
17. Allen P, Riordan PB, Hanrahan TJ, Joseph RL. Production and quality of boar and castrate bacon. I. Pig production, in-factory processing and carcass value. Ir J Food Tech.1981;5:93-104.
18. Wood JD, Riley JE. Comparison of boars and castrates for bacon production. 1. Growth data, and carcass and joint composition. Anim Prod.1982;35:55-63.
19. Nicholls LL, Price MA. A comparison of boars and barrows for growth and carcass characteristics at four slaughter weights. Agric Forestry Bulletin, Univ of Alberta, Special Issue 65th Annual Feeders' Day Rep.1986;120-122.
20. Newell JA, Boland JP. Performance, carcass composition, and fat composition of boars, gilts and barrows fed two levels of protein. Can J Anim Sci.1972;52:543-551.
21. Pay MG, Davies TE. Growth, food conversion, and carcass characteristics in castrated and entire male pigs fed three different dietary protein levels. J Agric Sci Camb.1973;81:65-68.
22. Fortin A, Friend DW, Sarkar NK. A note on the carcass composition of Yorkshire boars and barrows. Can J Anim Sci.1983;63:711-714.
23. Castell AG, Strain JH. Influence of diet and sex-type (boar, castrate or gilt) on live and carcass measurements of self-fed pigs from two breed lines differing in growth rates. Can J Anim Sci.1985; 65:185-195.
24. Knudson BK, Hogberg MG, Merkel RA, Allen RE, Magee WT. Developmental comparisons of boars and barrows: I. Growth rate, carcass and muscle characteristics. J Anim Sci.1985;61:789- 796.
25. Friend DW, Fortin A, Butler G, Poste LM, Kramer JKG, Burrow VD. Naked oats (Avena nuda) with and without lysine supplementation, for boars and barrows: Growth, carcass and meat quality, energy and nitrogen metabolism. Can J Anim Sci.1989;69:765-778.
26. Castell AG, Cliplef RL, McKay RM. Effects of diet, litter, and sex type on the performance (from 22 to 90 kg liveweight) and carcass measurements of crossbred pigs. Can J Anim Sci.1985;65:821-834.
27. Squires EJ, Adeola O, Young G, Hacker RR. The role of growth hormones, b-adrenergic agents and intact males in pork production: A review. Can J Anim Sci.1993;73:1-23.
28. Xue JL, Dial GD, Schuiteman J, Kramer A, Fisher C, Marsh WE, Morrison RB, Squires JE. Evaluation of growth, carcass, and compound concentrations related to boar taint in boars and barrows. Swine Hlth Prod.1995;3:155-160.
29. National Research Council. Nutrient Requirements of Swine. National Academy Press, 1988; Washington, D.C. 1988.
30. Prescott JHD, Lamming GE. The influence of castration on the growth of male pigs in relation to high levels of dietary protein. Anim Prod.1967;9:535-545.
31. Creswell DC, Wallace HD, Combs GE, Palmer AZ, West RL. Lysine and tryptophan in diets for boars and barrows. J Anim Sci.1975;40:167 (Abstr).
32. Luce WG, Johnson RK, Walters LE. Effects of levels of crude protein on performance of growing boars. J Anim Sci.1976;42:1207-1210.
33. Reinhard MK, Mahan DC, Workman BL, Cline JH, Fetter AW, Grito AP Jr. Effect of increasing dietary protein level, calcium and phosphorus on feedlot performance, bone mineralization and serum mineral values with growing swine. J Anim Sci.1976;43:770-780.
34. Taverner MR, Campbell RG, King RH. The relative protein and energy requirements of boars, gilts and barrows. Austr J Exp Agric Anim Husb.1977;17:574-580.
35. Tyler RW, Luce WG, Johnson RK, Maxwell CV, Hintz RL, Walters LE. The effects of level of crude protein on performance of growing boars. J Anim Sci.1983;57:364-372.
36. Williams WD, Cromwell GL, Stahly TS, Overfield JR. The lysine requirement of the growing boar versus barrow. J Anim Sci.1984;58:657-665.
37. Yen HT, Cole DJA, Lewis D. Amino acid requirements of growing pigs. 7. The response of pigs from 25 to 55 kg liveweight to dietary ideal protein. Anim Prod.1986;43:141-154.
38. Yen HT, Cole DJA, Lewis D. Amino acid requirements of growing pigs. 8. The response of pigs from 50 to 90 kg liveweight to dietary ideal protein. Anim Prod.1986;43:155-165.
39. Cooke R, Lodge GS, Lewis D. Influence of energy and protein concentration in the diet on the performance of growing pigs. 3. Response to differences in levels of both energy and protein. Anim Prod.1972;14:219-228.
40. Holmes CW, Carr JR, Pearson G. Some aspects of the energy and nitrogen metabolism of boars, gilts and barrows given diets containing different concentrations of protein. Anim Prod.1980;31:279- 289.
41. Just A. The net energy value of crude (catabolized) protein for growth in pigs. Livest Prod Sci. 1982;9:349-360.
42. Henry Y, Colléaux Y, Sève B. Effects of dietary level of lysine and of level and source of protein on feed intake, growth performance, and plasma amino acid pattern in the finishing pig. J Anim Sci. 1992;70:188-195.
43. Adeola O, Young LG. Dietary protein-induced changes in porcine muscle respiration, protein synthesis and adipose tissue metabolism. J Anim Sci.1989;67:664-673.
44. Sugahara M, Baker DH, Harmon BG, Jensen AH. Effect of excess levels of dietary crude protein on carcass development in swine. J Anim Sci.1969;29:598-601.
45. Pond WG, Varel VH, Dickson JS, Haschek WM. Comparative response of swine and rats to high-fiber or high-protein diets. J Anim Sci.1989;67:716-723.
46. Fuller MF, Cadenhead A, Mollison G, Sève. Effects of the amount and quality of dietary protein on nitrogen metabolism and heat production in growing pigs. Br J Nutr.1987;58:277-285.
47. Campbell RG, Taverner MR, Curic DM. The effects of sex and liveweight on the growing pig's response to dietary protein. Anim Prod.1988;46:123-130.
48. Campbell RG, Taverner MR. Genotype and sex effects on the relationship between energy intake and protein deposition in growing pigs. J Anim Sci.1988;66:676-686.
49. Kroeske D. Value of boars in fattening compared to hogs and gilts. Report at the Commission on Pig Production, EAAP-Meeting. Dublin, 1968.
50. Walstra P. Experiments in the Netherlands on the effect of castration of pigs in relation to feeding level. Meat production from entire male animals. DN Rhodes, J & A Churchill Ltd, London. 1969;p129-141.
51. Skjaerlund DM, Mulvaney DR, Bergen WG, Merkel RA. Skeletal muscle growth and protein turnover in neonatal boars and barrows. J Anim Sci.1994;72:315-321.
52. Charette LA. The effects of sex and age of male at castration on growth and carcass quality of Yorkshire swine. Can J Anim Sci. 1961;41:30-39.
53. Bates RO, Zumbrunnen C, Jesse GW. Effect of age at castration on swine. J Anim Sci.1992; 70(Suppl 1):49 (Abstr).
54. Mulvaney DR. Effects of castration and administration of androgens to castrated male pigs upon growth and carcass composition. PhD Dissertation.1984;Michigan State University, East Lansing.
55. Hansson I. Effect of sex and weight on growth, feed efficiency and carcass characteristics of pigs. 1. Growth rate and feed efficiency of boars, barrows and gilts. Swed J Agric Res.1974:209-218.
56. Hansen BC, Lewis AJ. Effects of dietary protein concentration (corn:soybean meal ratio) on nitrogen balance of growing boars, barrows, and gilts: mathematical descriptions. J Anim Sci.1993;71:2110-2121.
57. Meunier-Salaun MC, Vantrimponte MN, Raab A, Dantzer R. Effect of floor area restriction upon performance, behavior and physiology of growing-finishing pigs. J Anim Sci.1987;64:1371-1377.
58. Simonsen HB. Behavior and distribution of fattening pigs in the multi-activity pen. Appl Anim Behav Sci.1990;27:311-324.
59. Booth WD. Endocrine and exocrine factors in the reproductive behavior of the pig. Symp Zool Soc London.1980;45:289-311.
60. Bonneau M, Desmoulin B. Evolution de la teneur en androsténone du tissu adipeux dorsal chez la porc mâle entier de type Large White: variation selon les conditions d'élevage. Reprod Nutr Dév. 1980;20:1429-1437.
61. Patterson RLS, Lightfoot AL. Effect of sex grouping during growth on 5b-androstenone development in boars at three commercial slaughter weights. Meat Sci.1984;10:253-263.
62. Froseth JA, Matin EL, Hill JK. Effects of limited feeding, sex and liveweight on porcines. J Anim Sci.1973;37:262(Abstr).
63. Hansen BC, Lewis AJ. Effects of dietary protein concentration (corn:soybean meal ratio) on the performance and carcass characteristics of growing boars, barrows, and gilts: mathematical descriptions. J Anim Sci.1993;71:2122-2132.
64. Hansson I, Lundström K, Malmfors B. Effect of sex and weight on growth, feed efficiency and carcass characteristics of pigs. 2. Carcass characteristics of boars, barrows and gilts, slaughtered at four different weights. Swed J Agric Res.1975;5:69-80.
65. Campbell RG, Taverner MR. Effects of strain and sex on protein and energy metabolism in growing pigs. In: PW Moe, HF Tyrrell, PJ Reynolds (Ed.) Energy Metabolism of Farm Animals. 1985;p78 Eur Assoc Anim Prod Publ No 32 Rowan and Littlefield, USA
66. Wilson GD, Burnside JE, Bray RW, Phillips PH, Grummer RH. Pork carcass value as affected by protein level and supplementation with aureomycin and vitamin B12. J Anim Sci.1953;12:291- 296.
67. Greeley MG, Meade RJ, Hanson LE, Nordstrom J. Energy and protein intakes by growing swine. II Effects on rate and efficiency of gain and on carcass characteristics. J Anim Sci.1964;23:816-822.
68. Sather AP, Jones SDM, Joyal S. Feedlot performance, carcass composition and pork quality from entire male and female Landrace and Large White market-weight pigs. Can J Anim Sci. 1991;71:29- 42.
69. Sather AP, Jones SDM, Robertson WM, Zawadski S. Sex effects on fat hardness mater reading of market weight pigs. Can J Anim Sci.1995;75:509-515.
70. Wood JD. Fat quality in pig meat - UK. In: JD Wood (ed), Meat Research Institute Special Report No.2. A Workshop in CEC Program of Coordination of Research on Animal Husbandry, Brussels.1984;p9-14.
71. Edwards SA, Wood JD, Moncrieff CB, Porter SJ. Comparison of the Duroc and Large White as terminal sire breeds and their effect on pigmeat quality. Anim Prod. 1992;54:289-297.
72. Dransfield E, Kempster ANJ. Incidence of soft fat in British pigs. Anim Prod. 1988;88:502(abstr).
73. Wood JD, Enser M. Comparison of boars and castrates for bacon production. Anim Prod. 1982;35:65-74.
74. Malmfors B, Lundström K, Hansson I. Fatty acid composition of porcine back fat and muscle lipids as affected by sex, weight and anatomical location. Swed J Agric Res.1978;8:25-38.
75. Barton-Gade PA. Meat and fat quality in boars, castrates and gilts. Livest Prod Sci.1987;16:187- 196.
76. Nicholls LL, Price MA. A comparison of boars and barrows for meat quality characteristics and steroid concentrations at four slaughter weights. Agric Forestry Bulletin, Univ of Alberta, Special Issue 66th Annual Feeders' Day Rep.1987;30-32.
77. Smithard RR, Smith WC, Ellis M. A note on the fatty acid composition of backfat from boars in comparison with gilts and barrows. Anim Prod.1980;31:217-219.
78. Buck SF. A comparison of pigs slaughtered at three different weights. I. Carcass quality and performance. J Agric Sci Camb.1963;60:19-26.
79. Emerson JA, Pearson AM, Hoefer JA, Magee WT, Bratzler LJ. Effect of slaughter weight upon the processing characteristics, quality and consumer acceptability of pork carcasses and cuts. J Anim Sci.1964;23:436-443.
80. Vold E, Moen RA. A note on the effect of castration upon the development of the skin in the pig. Anim Prod.1972;14:253-254.
81. Knudson BK, Hogberg MG, Merkel RA, Allen RE, Magee WT. Developmental comparisons of boars and barrows: II. Body composition and bone development. J Anim Sci.1985;61:797-801.
82. Lundström K, Nilsson H, Malmfors B. Interrelationships between meat quality characteristics in pigs. Acta Agric Scand Supp.1979;l 21:71-80.
83. Warriss PD. The relationship between pH45 and drip in pig muscle. J Food Tech.1982;17:573- 578.
84. Tarrant PV, Gallwey WJ, McGloughlin P. Carcass pH values in Irish Landrace and Large White pigs. Ir J Agric Res.1979;18:167-172.
85. Moss BW, Robb JD. The effect of pre-slaughter lairage on serum thyroxine and cortisol levels at slaughter, and meat quality of boars, hogs and gilts. J Sci Food Agric.1978;29:689-696.
86. Ellis M, Smith WC, Clark JBK, Innes N. A comparison of boars, gilts and castrates for bacon manufacture. 1. On farm performance, carcass and meat quality characteristics and weight loss in the preparation of sides for curing. Anim Prod.1983;37:1-9.
87. Rose M, Adams D. Evidence for pain and suffering in other animals. In: Langley G (ed.) Animal Experimentation. Chapman & Hall, New York.1989.
88. Jackson WT. On-farm animal welfare law in Europe - using the law. Appl Anim Behav Sci. 1988;20:165-173.
89. McMahon, K. Iowa manure bill has 'something for everyone to hate'. National Hog Farmer. 1996;41(No.1):12-13.
90. McMahon, K. North Carolina tightens manure regulations. National Hog Farmer.1996; 41(No.4):28.
91. Anonymous. Paper on acidification. Ministry of Housing, Physical Planning and Environment/Ministry of Agriculture and Fisheries, The Hague, the Netherlands.1984;p59.
92. Anonymous. Bekendt gørelse om husdyrgødning og ensilage m.v. Miljoministeriets bekendtgørelse.1988;nr.568.
93. Sawyer GJ, Barker DJ. Growth promotants in cattle in Australia. Austr Vet J.1988;65:101-108.
94. Hancock DL, Wagner JF, Anderson DB. Effects of estrogens and androgens on animal growth. In Pearson AM, Dutson TR (ed), Advances in Meat Research. Vol 7 Growth Regulation in Farm Animals. Elservier Applied Science Publisher, Essex, UK.1991;p255-297.
95. Woehling HL, Wilson GD, Grummer RW, Bray RW, Casida LE. Effects of stilboestrol and testosterone pellets implanted into growing-fattening pigs. J Anim Sci.1951;10:889-892.
96. Pearson AM, Combs GE, Wallace HD, Sleeth RB, Stroud JW, Shepherd JM, Koger M. The effects of stilboestrol implants on swine of different sexes. J Anim Sci.1952;11:251-260.
97. Sleeth RB, Pearson AM, Wallace HD, Voropf DH, Koger M. Effect of injection of testosterone, estradiol and a combination of the two upon growing-fattening swine. J Anim Sci.1953;12:322-330.
98. Bidner TD, Merkel RA, Miller ER. Effect of a combination of diethylstilbestrol and methyltestosterone on performance, carcass traits, serum and muscle characteristics of pigs. J Anim Sci.1972;35:525-533.
99. Lucas LE, Peo ER, Jr, Svajgr AJ. Effect of a combination of diethylstilbestrol and methyltestosterone and protein level in the diet on performance and carcass traits in an unselected line of swine. J Anim Sci.1973;36:1094-1098.
100. DeWilde RO, Lauwer H. The effect of parenteral use of estradiol, progesterone, testosterone and trenbolone on growth and carcass composition in pigs. J Anim Sci.1984;59:1501-1509.
101. Denzer LE, Thompson LH, McKeith FK, Parrett DF, Thomas DL. Evaluation of growth, carcass traits and reproductive organs of young boars in response to zeranol implantation. J Anim Sci.1986;62:1164-1171.
102. Kluber EF, III, Minton JE, Stevenson JS, Hunt MC, Davis DL, Hoagland TA, Nelssen JL. Growth, carcass traits, boar odor and testicular and endocrine functions of male pigs fed a progestogen, altrenogest. J Anim Sci.1988;66:470-478.
103. Sheridan PJ, Austin FH, Bourke S, Roche JF. The effect of anabolic agents on growth rate and reproductive organs of pigs. Livest Prod Sci.1990;26:263-275.
104. Martinez M, López-Bote C, Sancho G, Ventanas J. Effects of trenbolone acetate on swine carcass characteristics and backfat composition. Can J Anim Sci.1992;72:969-972.
105. Hancock DL, Wagner JF, Anderson DB. Effects of estrogens and androgens on animal growth. In Pearson AM, Dutson TR, eds. Advances in Meat Research. Volume 7: Growth Regulation in Farm Animals. Elsevier Applied Science Publishers: Essex, England. 1991. 255-297.